Directed Improvement of Hybrid Rice Zhuoliangyou 1126 by Heavy Ion Beam Mutagenesis Based on M1TDS Targeted Screening Technology
SHAO Ye1,2,#, HU Yuanyi1,2,#, PENG Yan1,2,#, MAO Bigang1,2, LIU Huimin1,2, TANG Chanjuan1, LEI Bin1,2, TANG Li1,2, YU Lixia3, LI Wenjian3, LUO Wuzhong3, LUO Zhibin1,2, YUAN Yuantao1,2, LI Yaokui1,2, ZHANG Dan1,2, ZHOU Libin3, BAI Lianyang,2,4,*, TANG Wenbang,1,2,*, ZHAO Bingran,1,2,*
1Hunan Hybrid Rice Research Center, Changsha 410125, China
2National Key Laboratory of Hybrid Rice, Changsha 410125, China
3National Laboratory of Heavy Ion Accelerator, Institute of Modern Physics, Chinese Academy of Sciences, Lanzhou 730000, China
4Hunan Academy of Agricultural Sciences, Changsha 410125, China
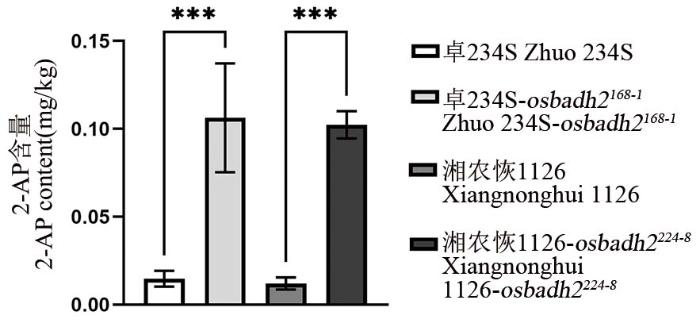
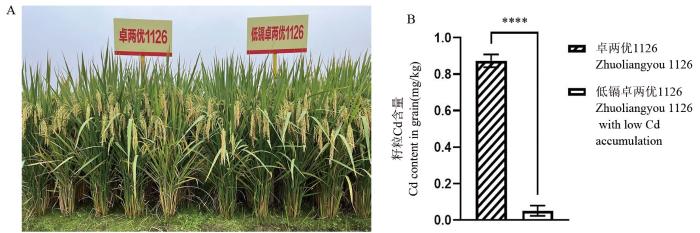
【Objective】Using the two-line hybrid rice Zhuoliangyou 1126 as the base variety, we aimed to rapidly develop upgraded varieties with traits such as low cadmium accumulation, fragrance, storage tolerance, submergence tolerance, and a low glycemic index.【Method】Zhuo 234S and Xiangnonghui 1126, the parental lines of Zhuoliangyou 1126, were mutagenized using heavy-ion beams. Chimeric mutants of the OsNRAMP5, OsBADH2, OsLOX3, OsPAO5, OsSSIIIa, and OsBEIIb genes were identified in the M1 generation via M1TDS technology. Mutations in the target genes were further detected and isolated in the M2 generation using Kompetitive Allele-Specific PCR (KASP) genotyping. Hybrid rice with low cadmium accumulation was developed by crossing parents carrying OsNRAMP5 mutations.【Results】A total of 13 chimeric mutants involving six target genes were identified in the M1 generation. Among these, seven were confirmed to carry homozygous or heterozygous mutations in the M2 generation. The cadmium content in OsNRAMP5 homozygous mutants was significantly lower than in the wild type, while the content of the aromatic compound 2-acetylpyrroline (2-AP) was significantly higher in OsBADH2 homozygous mutants. When the low-cadmium hybrid rice derived from OsNRAMP5-mutated parents was pot-grown in cadmium-contaminated soil (available Cd: 0.677 mg/kg, pH 5.6), the cadmium content in grains remained consistently below 0.05 mg/kg, compared to 0.91 mg/kg in the wild-type control.【Conclusion】The combination of heavy-ion beam mutagenesis and M1TDS technology enabled the rapid improvement of traits such as low cadmium accumulation in Zhuoliangyou 1126. This study provides a successful example and a general technical reference for transitioning from traditional mutagenesis breeding to directed mutagenesis breeding.
Keywords:rice;
heavy ion beam mutation breeding;
low cadmium accumulation;
fragrance
SHAO Ye, HU Yuanyi, PENG Yan, MAO Bigang, LIU Huimin, TANG Chanjuan, LEI Bin, TANG Li, YU Lixia, LI Wenjian, LUO Wuzhong, LUO Zhibin, YUAN Yuantao, LI Yaokui, ZHANG Dan, ZHOU Libin, BAI Lianyang, TANG Wenbang, ZHAO Bingran. Directed Improvement of Hybrid Rice Zhuoliangyou 1126 by Heavy Ion Beam Mutagenesis Based on M1TDS Targeted Screening Technology[J]. Chinese Journal of Rice Science, 2025, 39(5): 624-634 DOI:10.16819/j.1001-7216.2025.240106
重离子束具有传能线密度(Linear Energy Transfer, LET)高,即重离子经过的径迹区域集中地沉积大量能量的特性,相比低LET的γ射线与X射线具有更多优势:1)相对生物学效应大,即能够以较小的剂量对染色体局部造成破坏,产生多为不易正确修复的DNA双链断裂及团簇损伤,高效诱发遗传突变[6-7];2)对遗传背景影响较小,重离子束仅破坏其径迹经过的染色体区域,对基因组的其他区域不产生直接影响[8-9]。基于以上特点,重离子束辐照诱变具有“单点育种”(One-point breeding)的现象[8-9]。目前国内外学者利用重离子束诱变已在作物[10-13]、观赏植物[14]、微生物[15-16]中创制了一系列突变新种质。
217-288E, Mixed pool of leaf samples from the eastern side of plants in line 217-288 of M1 population; 217-288W, Mixed pool of leaf samples from the western side of plants in lines 217-288 of M1 population. And so on, for other number of mixed pools.
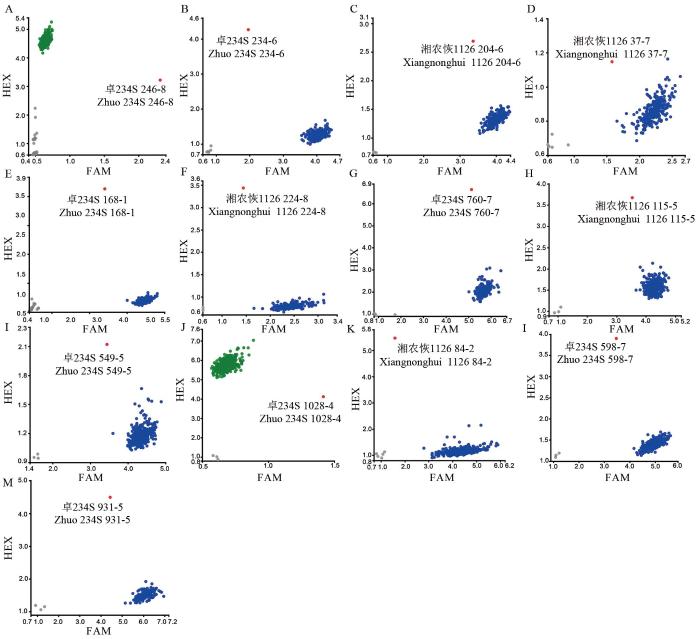
A~D分别代表OsNRAMP5基因不同突变位点基因分型;E, F 分别代表OsBADH2基因不同突变位点基因分型;G, H分别代表OsLOX3基因不同突变位点基因分型;I~K分别代表OsPAO5基因不同突变位点基因分型;L, M分别代表OsSSIIIa基因和OsBEIIb基因突变位点基因分型;A. 图中246-8代表诱变M1群体246行第8株(下同);蓝点/绿点代表野生基因型,红点代表突变基因型,灰点代表阴性对照;FAM和HEX分别代表两种不同颜色的荧光修饰标签。
Fig. 1.
KASP genotyping in the mixed pool of target gene mutations
A-D represent the genotyping of different mutation sites of OsNRAMP5 gene; E and F represent the genotyping of different mutation sites of OsBADH2 gene, respectively; G and H represent genotyping of different mutation sites of OsLOX3 gene, respectively; I-K represent genotyping of different mutation sites of OsPAO5 gene; L, M represent the genotyping of mutation sites in OsSSIIIa gene and OsBEIIb gene, respectively; In panel A, 246-8 represents the 8th plant in 246 rows of M1 population (the same below); Blue/green dots represent wild genotypes, red dots represent mutant genotypes, and gray dots represent negative controls; FAM and HEX represent fluorescently modified labels of two different colors, respectively.
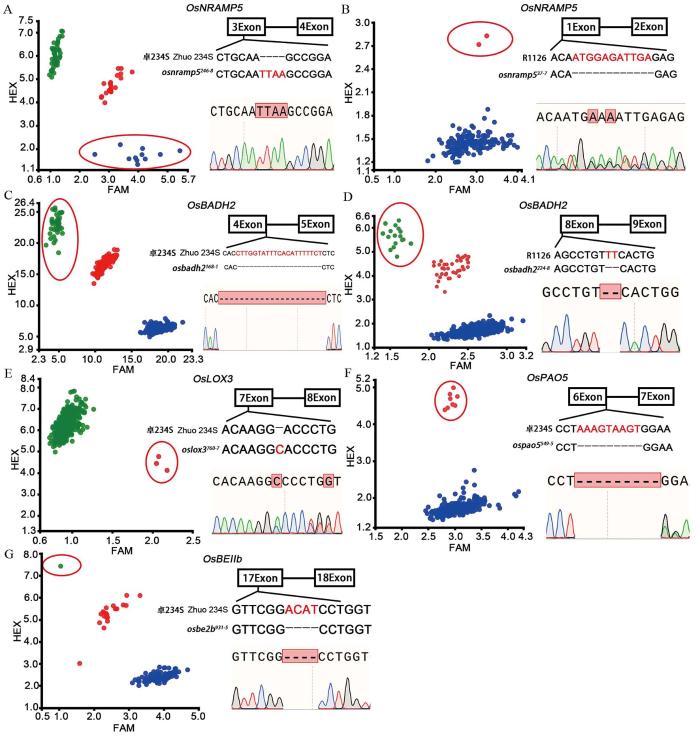
A, B:卓234S和湘农恢1126(R1126) M2群体OsNRAMP5基因KASP鉴定及突变单株Sanger测序验证,osnramp5246-8和osnramp537-7分别代表卓234S和湘农恢1126的OsNRAMP5基因突变单株; C, D:卓234S和湘农恢1126 M2群体OsBADH2基因KASP鉴定及Sanger测序验证,osbadh2168-1和 osbadh2224-8分别代表卓234S和湘农恢1126的OsBADH2基因突变单株;E~G:卓234S M2群体OsLOX3、OsPAO5和OsBEⅡb基因KASP鉴定及Sanger测序验证,oslox3760-7、ospao5549-5和osbe2b931-5分别代表卓234S的OsLOX3、OsPAO5和OsBEⅡb基因突变单株。图A中红点代表杂合基因型,蓝点代表纯合突变基因型,绿点代表野生基因型;图B~G中红点代表杂合基因型,绿点代表纯合突变基因型,蓝点代表野生基因型;红色椭圆形框内圆点代表目标基因纯合/杂合突变基因型。
Fig. 2.
KASP identification of M2 generation and Sanger sequencing of mutant plants
A and B, KASP identification of OsNRAMP5 gene in Zhuo 234S and Xiangnonghui 1126 M2 population and Sanger sequencing of mutants. osnramp5246-8 and osnramp537-7 represent mutant plant of OsNRAMP5 gene in Zhuo 234S and Xiangnonghui 1126, respectively; C and D, KASP identification of OsBADH2 gene in Zhuo 234S and Xiangnonghui 1126 M2 population and Sanger sequencing of mutants; osbadh2168-1and osbadh2224-8 represent mutant plant of OsBADH2 gene in Zhuo234S and Xiang Nonghui 1126, respectively. E-G, KASP identification of OsLOX3, OsPAO5 and OsBEⅡb gene in Zhuo 234S M2 population and Sanger sequencing of mutants. oslox3760-7, ospao5549-5 and osbe2b931-5 represented mutant plant of OsLOX3, OsPAO5 and OsBEⅡb gene of Zhuo 234S, respectively. In panel A, red dots represent heterozygous genotype, blue dots represent homozygous mutant genotype, green dots represent wild genotype. In panels B-G, red dots represent heterozygous genotype, green dots represent homozygous mutant genotype, blue dots represent wild genotype. The dots in the red oval box represent the homozygous/heterozygous genotype of target genes.
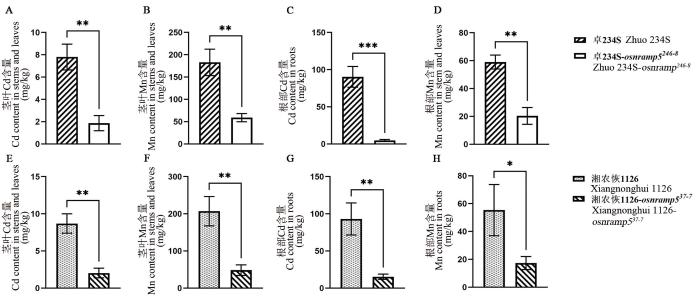
Fig. 3.
Contents of Cd and Mn in homozygous mutant of OsNRAMP5 and control
A-D, Contents of Cd and Mn in homozygous mutant Zhuo 234S-osnramp5246-8 and its original control material; E-H, Contents of Cd and Mn in the homozygous mutant Xiangnonghui 1126-osnramp537-7and its original control material; *, **, and *** represent difference was significant at 0.05, 0.01, and 0.001 levels, respectively (t-test).
Studies on biological effects of ion beams on lethality, molecular nature of mutation, mutation rate, and spectrum of mutation phenotype for mutation breeding in higher plants
[J]. Journal of Radiation Research, 2010, 51(3): 223-233.
Targeted mutagenesis of POLYAMINE OXIDASE 5 that negatively regulates mesocotyl elongation enables the generation of direct-seeding rice with improved grain yield
Role of isolated and clustered DNA damage and the post-irradiating repair process in the effects of heavy ion beam irradiation
1
2015
... 重离子束具有传能线密度(Linear Energy Transfer, LET)高,即重离子经过的径迹区域集中地沉积大量能量的特性,相比低LET的γ射线与X射线具有更多优势:1)相对生物学效应大,即能够以较小的剂量对染色体局部造成破坏,产生多为不易正确修复的DNA双链断裂及团簇损伤,高效诱发遗传突变[6-7];2)对遗传背景影响较小,重离子束仅破坏其径迹经过的染色体区域,对基因组的其他区域不产生直接影响[8-9].基于以上特点,重离子束辐照诱变具有“单点育种”(One-point breeding)的现象[8-9].目前国内外学者利用重离子束诱变已在作物[10-13]、观赏植物[14]、微生物[15-16]中创制了一系列突变新种质. ...
Recent advances in the biology of heavy-ion cancer therapy
1
2010
... 重离子束具有传能线密度(Linear Energy Transfer, LET)高,即重离子经过的径迹区域集中地沉积大量能量的特性,相比低LET的γ射线与X射线具有更多优势:1)相对生物学效应大,即能够以较小的剂量对染色体局部造成破坏,产生多为不易正确修复的DNA双链断裂及团簇损伤,高效诱发遗传突变[6-7];2)对遗传背景影响较小,重离子束仅破坏其径迹经过的染色体区域,对基因组的其他区域不产生直接影响[8-9].基于以上特点,重离子束辐照诱变具有“单点育种”(One-point breeding)的现象[8-9].目前国内外学者利用重离子束诱变已在作物[10-13]、观赏植物[14]、微生物[15-16]中创制了一系列突变新种质. ...
Studies on biological effects of ion beams on lethality, molecular nature of mutation, mutation rate, and spectrum of mutation phenotype for mutation breeding in higher plants
3
2010
... 重离子束具有传能线密度(Linear Energy Transfer, LET)高,即重离子经过的径迹区域集中地沉积大量能量的特性,相比低LET的γ射线与X射线具有更多优势:1)相对生物学效应大,即能够以较小的剂量对染色体局部造成破坏,产生多为不易正确修复的DNA双链断裂及团簇损伤,高效诱发遗传突变[6-7];2)对遗传背景影响较小,重离子束仅破坏其径迹经过的染色体区域,对基因组的其他区域不产生直接影响[8-9].基于以上特点,重离子束辐照诱变具有“单点育种”(One-point breeding)的现象[8-9].目前国内外学者利用重离子束诱变已在作物[10-13]、观赏植物[14]、微生物[15-16]中创制了一系列突变新种质. ...
Calibration-free NGS quantitation of mutations below 0.01% VAF
1
2021
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Selective multiplexed enrichment for the detection and quantitation of low-fraction DNA variants via low-depth sequencing
1
2021
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield
1
2017
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Production of aromatic three-line hybrid rice using novel alleles of BADH2
1
2022
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Antisense suppression of LOX3 gene expression in rice endosperm enhances seed longevity
1
2015
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Targeted mutagenesis of POLYAMINE OXIDASE 5 that negatively regulates mesocotyl elongation enables the generation of direct-seeding rice with improved grain yield
1
2021
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes
1
2017
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Loss of function of SSIIIa and SSIIIb coordinately confers high RS content in cooked rice
1
2023
... 最近发展起来的肿瘤无创早期诊断液体活检技术,通过深度靶向测序实现了血液中肿瘤的低频嵌合突变检测,50 000×的平均深度能准确检出0.1%的低频突变[23-24].受此启发,赵炳然团队前期发明了“一种高通量靶向鉴定理化诱变植株M1代突变及获取突变体的方法”(简称M1TDS技术:M1 generation targeted deep sequencing technology,专利号ZL201911223356.0).该技术突破了传统理化诱变育种难以高效定向获取目标基因突变的技术瓶颈,利用其超强的低频突变鉴定能力,在水稻理化诱变M1代即实现了目标基因突变的检出,快速创制出遗传背景、综合农艺性状与原始品种臻两优8612无明显差异,而在中、重度镉污染田种植表现镉低积累特性的杂交稻“莲两优1号”(即镉低积累臻两优8612)等[12].在此基础上,本研究以两系杂交水稻卓两优1126父母本为材料,通过重离子束辐射和M1TDS技术靶向筛选,创制出镉低积累(OsNRAMP5)[25]、香味(OsBADH2)[26]、耐储藏(OsLOX3)[27]、适宜直播(OsPAO5)[28]、低升糖指数(OsSSIIIa、OsBEIIb)[29-30]等有重要利用价值的水稻新种质,在不到2年的时间内就研发出与原始材料遗传背景、综合农艺性状均没有明显差异的镉低积累杂交稻组合卓两优1126,为在各种生物中开展重离子束辐射定向遗传改良提供了技术参考. ...
Detection and characterization of genome-wide mutations in M1 vegetative cells of gamma-irradiated Arabidopsis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}