A map of rice genome variation reveals the origin of cultivated rice
1
2012
... 亚洲栽培稻(Oryza sativa L.)是全球三大主要粮食作物之一,是世界一半以上、中国六成人口的主粮[1].在过去近百年的时间里,得益于无数育种专家的不懈努力,中国水稻育种取得了巨大的进步,经历了三个重要的发展阶段:矮化育种、杂交育种以及绿色超级稻的培育,实现了三次重大的飞跃[2].水稻平均产量显著提升,从1950年的140.5 kg/亩增加至2022年的472 kg/亩(数据来源:国家统计局). ...
中国水稻遗传育种历程与展望
2
2018
... 亚洲栽培稻(Oryza sativa L.)是全球三大主要粮食作物之一,是世界一半以上、中国六成人口的主粮[1].在过去近百年的时间里,得益于无数育种专家的不懈努力,中国水稻育种取得了巨大的进步,经历了三个重要的发展阶段:矮化育种、杂交育种以及绿色超级稻的培育,实现了三次重大的飞跃[2].水稻平均产量显著提升,从1950年的140.5 kg/亩增加至2022年的472 kg/亩(数据来源:国家统计局). ...
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
The history and prospect of rice genetic breeding in China
2
2018
... 亚洲栽培稻(Oryza sativa L.)是全球三大主要粮食作物之一,是世界一半以上、中国六成人口的主粮[1].在过去近百年的时间里,得益于无数育种专家的不懈努力,中国水稻育种取得了巨大的进步,经历了三个重要的发展阶段:矮化育种、杂交育种以及绿色超级稻的培育,实现了三次重大的飞跃[2].水稻平均产量显著提升,从1950年的140.5 kg/亩增加至2022年的472 kg/亩(数据来源:国家统计局). ...
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
Genomic architecture of heterosis for yield traits in rice
1
2016
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
Hybrid rice technology for food security in the world
2
2004
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
Hybrid vigor in rice
1
1926
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
水稻的雄性不孕性
1
1966
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
A Preliminary report on male sterility in rice, Oryza sativa L
1
1966
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
中国杂交水稻推广历程及现状分析
4
2016
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... [7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
Review of extension and analysis on current status of hybrid rice in China
4
2016
... 杂种优势(heterosis)是指杂交F1代表现出各性状优于亲本的特性,具体表现为产量、品质、适应性和抗病虫害能力的提升[3].杂交育种被认为是提高作物产量最有效的途径[4].1926年,Jones等[5]报道了水稻的杂种优势.中国杂交水稻的研究始于1966年袁隆平对雄性不育现象的报道[6].1970年,袁隆平的助手李必湖在海南发现了一株天然细胞质雄性不育株,并命名为“野败”;1973年,张先程从东南亚品种中找到了可以有效恢复野败育性的恢复系;1973年10月,袁隆平在苏州召开的水稻科研会议上发表了题为《利用野败选育三系的进展》的论文,标志着中国籼型杂交水稻三系配套成功[2].1976年,三系杂交水稻开始在全国大面积推广,杂种优势的利用使中国水稻进入第二次绿色革命.1995年,杂交稻种植面积在全国水稻种植面积占比达到历史最高点(68%)[7].杂交水稻为中国粮食安全做出了巨大的贡献. ...
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... [7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
Male sterility and fertility restoration in crops
3
2014
... 三系法杂交水稻由雄性不育系、保持系和恢复系组成.其中,细胞质型雄性不育系(Cytoplasmic Male Sterile)因线粒体缺陷而导致花粉败育,保持系与雄性不育系基因组相同,但线粒体无缺陷,恢复系含有恢复基因,可以恢复雄性不育系的育性[8].恢复系和雄性不育系分别作为父母本,用于生产商业化的杂交种.保持系与雄性不育系杂交后,其后代依然保持雄性不育的特性,这使得它们可以用于次年的杂交制种过程[8].三系杂交系统已被广泛应用了将近50年,但具有恢复基因的水稻极少,可作为父本水稻的选择性狭窄,在一定程度上限制了三系法在生产中的应用[9]. ...
... [8].三系杂交系统已被广泛应用了将近50年,但具有恢复基因的水稻极少,可作为父本水稻的选择性狭窄,在一定程度上限制了三系法在生产中的应用[9]. ...
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
杂交水稻发展的战略
1
2018
... 三系法杂交水稻由雄性不育系、保持系和恢复系组成.其中,细胞质型雄性不育系(Cytoplasmic Male Sterile)因线粒体缺陷而导致花粉败育,保持系与雄性不育系基因组相同,但线粒体无缺陷,恢复系含有恢复基因,可以恢复雄性不育系的育性[8].恢复系和雄性不育系分别作为父母本,用于生产商业化的杂交种.保持系与雄性不育系杂交后,其后代依然保持雄性不育的特性,这使得它们可以用于次年的杂交制种过程[8].三系杂交系统已被广泛应用了将近50年,但具有恢复基因的水稻极少,可作为父本水稻的选择性狭窄,在一定程度上限制了三系法在生产中的应用[9]. ...
The strategy for hybrid rice development
1
2018
... 三系法杂交水稻由雄性不育系、保持系和恢复系组成.其中,细胞质型雄性不育系(Cytoplasmic Male Sterile)因线粒体缺陷而导致花粉败育,保持系与雄性不育系基因组相同,但线粒体无缺陷,恢复系含有恢复基因,可以恢复雄性不育系的育性[8].恢复系和雄性不育系分别作为父母本,用于生产商业化的杂交种.保持系与雄性不育系杂交后,其后代依然保持雄性不育的特性,这使得它们可以用于次年的杂交制种过程[8].三系杂交系统已被广泛应用了将近50年,但具有恢复基因的水稻极少,可作为父本水稻的选择性狭窄,在一定程度上限制了三系法在生产中的应用[9]. ...
作物育性调控和分子设计杂交育种前沿进展与展望
1
2021
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
Fertility regulation and molecular design hybrid breeding in crops
1
2021
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
对光照长度敏感的隐性雄性不育水稻的发现与初步研究
2
1985
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
... 1973年,石明松在湖北省沔阳沙湖原种场的农垦58群体中发现了一株天然雄性不育水稻,当地的35个品种与其杂交产生的F1代结实率均保持在80%以上,在F2代群体中,可育与不育的分离比接近3∶1,说明该不育表型受一对隐性核基因控制,中国农业科学院邓景扬研究员将该基因命名为“光感核不育基因”[11].籼型温敏型雄性不育株系培矮64S是由农垦58S转育而来,但随着遗传背景从粳稻转移至籼稻,其育性转换变得更依赖于温度变化而非光照时长. ...
The discovery and study of the photosensitive recesseive male-sterile rice
2
1985
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
... 1973年,石明松在湖北省沔阳沙湖原种场的农垦58群体中发现了一株天然雄性不育水稻,当地的35个品种与其杂交产生的F1代结实率均保持在80%以上,在F2代群体中,可育与不育的分离比接近3∶1,说明该不育表型受一对隐性核基因控制,中国农业科学院邓景扬研究员将该基因命名为“光感核不育基因”[11].籼型温敏型雄性不育株系培矮64S是由农垦58S转育而来,但随着遗传背景从粳稻转移至籼稻,其育性转换变得更依赖于温度变化而非光照时长. ...
晚粳自然两用系选育及应用初报
1
1981
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
A perliminary report on selection and application of natural dual-purpose lines of late japonica
1
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
中国两系杂交水稻选育与应用进展
2
2009
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
Progress of application and breeding on two-line hybrid rice in China
2
2009
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
两系法杂交水稻研究的进展
1
1990
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
Research progress on two-line hybrid rice
1
1990
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
我国两系杂交水稻发展的现状和建议
1
2011
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
Current situation and suggestions for development of two-line hybrid rice in China
1
2011
... 细胞核雄性不育(Genic Male Sterile, GMS)由核基因组控制.许多调控减数分裂、绒毡层或雄配子发育的基因一旦发生突变,很容易产生雄性不育.绝大多数核雄性不育材料为完全雄性不育,难以进行纯合不育系的繁衍,因此无法用于杂交制种[10].有一部分核不育系的育性受到环境条件影响,称为环境敏感型核雄性不育系(Environment-sensitive Genic Male Sterile, EGMS),其中,光/温敏型核雄性不育系(Photoperiod/Thermo-sensitive Genic Male Sterile, P/TGMS)可以在高温或长日照条件下作为雄性不育系,也可以在低温或短日照条件下表现为可育并进行自我繁殖[8].因为P/TGMS系的不育基因受到核基因隐性控制,所以绝大多数的品系都可以作为它的恢复系.1973年,石明松先生在湖北首次发现了光敏感型雄性不育水稻,将其命名为“农垦58S”[11].1981年,石明松提出基于农垦58S的特性,通过“两系法”利用杂种优势[12].1994年,从农垦58S转育的一系列不育系配组育成的两系杂交组合通过省级审定,标志着中国两系杂交水稻正式进入生产应用阶段[13].1995年8月,在湖南怀化召开的两系法杂交中稻现场会上,袁隆平宣布两系杂交水稻取得成功,开始大面积推广[7].两系杂交系统是杂交水稻的一项重要创新,并成为中国大规模粮食生产中更受欢迎的方式[14,15].2013年,“两系法杂交水稻技术研究与应用”获得了国家科学技术进步奖特等奖. ...
Workable male sterility systems for hybrid rice: Genetics, biochemistry, molecular biology, and utilization
1
2014
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
两系法杂交水稻的育种成就与展望
2
2021
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... [17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
Achievements and prospects of two-line system hybrid rice breeding
2
2021
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... [17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
2
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... [18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
2
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... [18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
长江中下游稻区籼型两系与三系杂交水稻组合的比较分析
2
2009
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
Comparison analysis between two-line and three-line indica hybrid rice combinations in the middle and down area of the Yangtse Valley of China
2
2009
... 从2002年开始,首批育成的两系超级杂交稻组合“两优培九”种植面积超过广泛应用的三系组合“汕优63”,成为当年推广面积最大的杂交稻[7].2009年开始,“扬两优6号”、“Y两优1号”等一批优势组合开始包揽杂交水稻种植面积的前三名[7].2012年,两系杂交水稻已经覆盖了约4950万亩的总种植面积,约占杂交水稻总种植面积的三分之一[16],2016年上升至约50%[17,18].根据国家水稻数据中心的统计,自2015年起,两系法国审水稻品种数量开始逐渐超过三系;2017年全国推广面积前十的杂交组合有6个是两系杂交稻,到2019年这个数字上升到了7个;2019—2021年,全国共育成两系水稻不育系403个,占育成的不育系总量的58.83%,育成两系杂交稻国审品种719个,占育成杂交稻品种的57.34%[17].在2023年发布的《国家农作物优良品种推广目录》中有10个骨干型水稻品种,三系和两系品种分别有2个和3个.三个两系杂交水稻品种分别为晶两优534、晶两优华占和隆两优华占.2017—2021年,这三个两系杂交品种累计推广面积达5967万亩,而两个三系杂交品种累计推广面积约1761万亩[18].两系杂交稻虽然起步比三系杂交稻晚了近20年,但至今无论从不育系还是杂交品种数量,都已超过了三系杂交稻[19]. ...
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
2020年我国审定的水稻品种基本特性分析
1
2021
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
Analysis on Characteristics of Rice Varieties Registered in China in 2020
1
2021
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
两系和三系杂交水稻产量性状差异性比较
1
2022
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
Comparison of difference in yield characters between two-line and three-line hybrid rice
1
2022
... 与三系育种相比,两系育种系统的P/TGMS株系具有种质资源广泛、配组自由、杂种优势强、育种生产流程简单等优点.近年来,两系杂交水稻已在中国大规模应用,现有组合的产量、抗性、米质均不同程度优于相应组别的三系杂交水稻或常规水稻对照品种[4,13].程本义等[19]对长江中下游稻区2005―2008年期间进行区试的籼型两系三系杂交组合进行了比较分析,发现两系组合总体上产量高于三系2%~3%,米质总体略优于三系,抗病虫性则与三系相当.林海等[20]对2015至2020间的国家审定品种区试产量比较中也发现两系优于三系的产量特征,在2020年通过审定的籼型两系杂交稻的平均产量为609.99 kg/亩,籼型三系杂交稻平均产量为589.82 kg/亩.唐世军等[21]在四川盆地对两系和三系杂交水稻进行田间试验,比较两者在有效穗、穗粒数、千粒重、单穗重及产量上的差异,发现两系杂交水稻的平均产量、有效穗数、结实率高于三系.两系杂交组合的米质总体上也好于三系,历年种植面积在100万亩以上并达到优质食用稻二级以上水平的15个杂交水稻组合中,两系杂交稻有9个(数据来源:《2019年中国水稻产业发展报告》). ...
超级杂交水稻育种研究的进展
1
2008
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
Research progress on super hybrid rice breeding
1
2008
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
中籼型优质光温敏核不育系广占63S的选育
2
2002
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
... 从1973年光温敏现象被发现以来,经过几十年的发展,人们对光温敏现象的理解正在逐步深入.我国的光温敏不育系审定品种数量不断增长,但这些不育系的基因来源却十分单一,绝大多数都受到pms3或tms5位点的控制.随着两系育种的不断发展,tms5逐渐成为了最主流的光温敏位点.两系法制种极其依赖天气,因此需要保证十分彻底的高温不育性和尽可能低的育性转换温度.2000年华中农业大学的人工气候箱育性鉴定结果表明tms5控制的广占63S在高温、长日照下的不育性比pms3控制的培矮64S更好[23],吴亚先等[77]研究了12份温敏不育株系的低温敏感性,发现隆科638S、晶4155S的不育起点温度低于或接近23℃,Y58S的不育起点温度达到了23.0~23.5℃,广占63-4S则高于23.5℃.这也是2005年以后培矮64S逐渐被广占63S取代,广占63S又逐渐被Y58S、隆科638S、晶4155S等品种相继取代,成为应用最广泛的不育系的重要原因之一. ...
Breeding of fine quality PTGMS line Guangzhan 63S in medium indica rice
2
2002
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
... 从1973年光温敏现象被发现以来,经过几十年的发展,人们对光温敏现象的理解正在逐步深入.我国的光温敏不育系审定品种数量不断增长,但这些不育系的基因来源却十分单一,绝大多数都受到pms3或tms5位点的控制.随着两系育种的不断发展,tms5逐渐成为了最主流的光温敏位点.两系法制种极其依赖天气,因此需要保证十分彻底的高温不育性和尽可能低的育性转换温度.2000年华中农业大学的人工气候箱育性鉴定结果表明tms5控制的广占63S在高温、长日照下的不育性比pms3控制的培矮64S更好[23],吴亚先等[77]研究了12份温敏不育株系的低温敏感性,发现隆科638S、晶4155S的不育起点温度低于或接近23℃,Y58S的不育起点温度达到了23.0~23.5℃,广占63-4S则高于23.5℃.这也是2005年以后培矮64S逐渐被广占63S取代,广占63S又逐渐被Y58S、隆科638S、晶4155S等品种相继取代,成为应用最广泛的不育系的重要原因之一. ...
水稻光温敏雄性核不育系的系谱分析
2
2012
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
... 中国生产应用的两系核不育系,绝大多数(>95%)由农垦58S、安农S-1、株1S等几种早年选育品种衍生而来[24].株1S是杨远柱从抗罗早///科辐红2号/HA79317-7//02428组合的F2群体中发现的,于1997年育成的偏籼型广亲和温敏不育系[50].从亲缘上看,株1S与安农S-1并无直接关系,但株1S及其衍生的不育系均携带有与安农S-1一样的tms5温敏位点[51].严维等通过全基因组多态性分析发现,株1S及其衍生系与安农S-1及其衍生系,在TMS5区域和上下游各1 Mbp范围内存在多个多态性SNP位点和InDel标记,说明二者在该功能区段的来源并不相同[52]. ...
Pedigree analysis of photoperiod-thermo sensitive genic male sterile rice
2
2012
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
... 中国生产应用的两系核不育系,绝大多数(>95%)由农垦58S、安农S-1、株1S等几种早年选育品种衍生而来[24].株1S是杨远柱从抗罗早///科辐红2号/HA79317-7//02428组合的F2群体中发现的,于1997年育成的偏籼型广亲和温敏不育系[50].从亲缘上看,株1S与安农S-1并无直接关系,但株1S及其衍生的不育系均携带有与安农S-1一样的tms5温敏位点[51].严维等通过全基因组多态性分析发现,株1S及其衍生系与安农S-1及其衍生系,在TMS5区域和上下游各1 Mbp范围内存在多个多态性SNP位点和InDel标记,说明二者在该功能区段的来源并不相同[52]. ...
广适性水稻光温敏不育系Y58S的选育
1
2005
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
Breeding of the PTGMS line Y58S with wide adaptability in rice
1
2005
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
优质抗病高配合力中籼两用核不育系隆科638S的选育及应用
1
2021
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
Breeding and application of TGMS line Longke 638S with blast-resistance, good-quality and high combining-ability in middle-season indica rice
1
2021
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
抗病优质高配合力中籼型两用核不育系晶4155S的选育及应用
1
2021
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
Breeding and application of medium indica TGMS line Jing 4155S with blast resistence, good grain quality and high conbining ability in rice
1
2021
... 光温敏核不育系是两系杂交水稻的核心.1994年开始,两系育种快速发展的二十多年时间里,育种家们先后培育出多个重要的光温敏核不育株系.衍生自农垦58S的籼稻品种培矮64S在两系杂交稻推广过程中发挥了重要作用,两优培九是江苏省农业科学院粮食作物研究所以培矮64S为母本,93-11(扬稻6号)为父本育成的超级杂交稻先锋品种,推广初期示范点平均产量超过700 kg/亩,对比全国推广面积第一的三系杂交稻汕优63增产5.1%,大规模推广至全国后,平均产量达到了550 kg/亩,远超同期全国水稻平均产量(420 kg/亩)和杂交水稻平均产量(470 kg/亩)[22].1999年,辽宁省农业科学院稻作研究所杨振玉利用广亲和不育系N422S和广东优质籼稻品种广占63杂交育成了优质光温敏核不育系“广占63S”[23],其衍生不育系广占63-4S、宣69S和新安S等在两系推广过程中都发挥了重大作用,其中广占63S与93-11配成的“丰两优1号”是2008年推广面积最大的杂交稻,广占63-4S与9311配成的“扬两优6号”的推广面积在2009年、2010年、2011年分别位于杂交稻中的第一、第三和第二[24].2003年,邓启云等[25]利用安农S-1作为不育基因供体,通过与常菲22B、Lemont杂交、双交,聚合优质、高光效、抗病、抗逆等优良性状,再通过与培矮64S复交导入优良株叶形态和高配合力实现有利多基因累加,育成广适性光温敏核不育系“Y58S”.其与93-11的组合品种“Y两优1号”在2004年的超级稻区试中产量名列第一,比两优培九增产11.2%,并从2011年开始取代扬两优6号成为全国推广面积最大的杂交稻.2010年到2012年,湖南亚华种业科学研究院杨远柱选择优质软米不育系湘陵628S作为母本,利用分子标记辅助选择,将Pia、Pita等抗稻瘟病优质基因聚合在一起,育成了隆科638S和晶4155S,成为近两年应用最广泛的不育系[26,27]. ...
A long noncoding RNA regulates photoperiod-sensitive male sterility, an essential component of hybrid rice
1
2012
... 2012年,张启发团队发现了农垦58S中PMS3的一个点突变导致了其光敏表型.PMS3转录产生一个1236 bp的长链非编码RNA,被称为长日照特异性雄性不育相关RNA(long-day-specific male-fertility-associated RNA, LDMAR).不育系中一个单碱基突变改变了其二级结构,导致启动子甲基化程度升高,转录水平下降,最终造成花药细胞的提前程序性死亡并雄性不育[28].同年,庄楚雄和刘耀光团队在培矮64S中定位到其温敏位点为非编码RNA基因P/TMS12-1,后续证实与农垦58S中的pms3为同一突变位点.该团队进一步研究发现P/TMS12-1转录产物会被切割成21核苷酸的非编码小RNA.该小RNA被命名为Osa-smR5864,在培矮64S中产生单碱基替换,推测该突变会引起功能缺失而导致高温长日照下不育[29]. ...
Photoperiod- and thermo-sensitive genic male sterility in rice are caused by a point mutation in a novel noncoding RNA that produces a small RNA
1
2012
... 2012年,张启发团队发现了农垦58S中PMS3的一个点突变导致了其光敏表型.PMS3转录产生一个1236 bp的长链非编码RNA,被称为长日照特异性雄性不育相关RNA(long-day-specific male-fertility-associated RNA, LDMAR).不育系中一个单碱基突变改变了其二级结构,导致启动子甲基化程度升高,转录水平下降,最终造成花药细胞的提前程序性死亡并雄性不育[28].同年,庄楚雄和刘耀光团队在培矮64S中定位到其温敏位点为非编码RNA基因P/TMS12-1,后续证实与农垦58S中的pms3为同一突变位点.该团队进一步研究发现P/TMS12-1转录产物会被切割成21核苷酸的非编码小RNA.该小RNA被命名为Osa-smR5864,在培矮64S中产生单碱基替换,推测该突变会引起功能缺失而导致高温长日照下不育[29]. ...
PMS1T, producing phased small-interfering RNAs, regulates photoperiod-sensitive male sterility in rice
1
2016
... 农垦58S的另一温敏位点pms1为不完全显性控制,位于7号染色体,其幼穗中的转录本在miR2118的作用下会被剪切,生成21个核苷酸的阶段性干扰小RNA(phased small-interfering RNA, phasiRNA).在农垦58S中,miR2118识别位点附近的一个单核苷酸多态性(SNP)导致了在长日照条件下phasiRNA的大量积累,最终通过其他未知机制导致了雄性不育[30]. ...
安农S-1的研究及其利用概况
1
1999
... 1987年,邓华凤在3个籼稻材料的三交组合(超40B/H285//6209-3)F5代群体中,发现了1株天然雄性不育株,后经不同生态条件下两代选择育成了籼型温敏型雄性不育株系安农S-1[31].2014年庄楚雄和曹晓风团队发现其温敏特性由tms5位点控制,TMS5编码的RNase ZS1是一种RNA酶,可以特异性地降解泛素-60S核糖体蛋白L40基因(ubiquitin-60S ribosomal protein L40 family, UbL40)的mRNA.在安农S-1中,TMS5基因的一个T到A的单碱基突变导致了该基因在第71个碱基对处提前终止.推测安农S-1中的RNase ZS1功能缺失,致使高温下UbL40的转录本过度积累,最终导致雄性不育[32]. ...
An overview of research and utilization of Annong S-1
1
1999
... 1987年,邓华凤在3个籼稻材料的三交组合(超40B/H285//6209-3)F5代群体中,发现了1株天然雄性不育株,后经不同生态条件下两代选择育成了籼型温敏型雄性不育株系安农S-1[31].2014年庄楚雄和曹晓风团队发现其温敏特性由tms5位点控制,TMS5编码的RNase ZS1是一种RNA酶,可以特异性地降解泛素-60S核糖体蛋白L40基因(ubiquitin-60S ribosomal protein L40 family, UbL40)的mRNA.在安农S-1中,TMS5基因的一个T到A的单碱基突变导致了该基因在第71个碱基对处提前终止.推测安农S-1中的RNase ZS1功能缺失,致使高温下UbL40的转录本过度积累,最终导致雄性不育[32]. ...
RNase Z(S1) processes UbL40 mRNAs and controls thermosensitive genic male sterility in rice
2
2014
... 1987年,邓华凤在3个籼稻材料的三交组合(超40B/H285//6209-3)F5代群体中,发现了1株天然雄性不育株,后经不同生态条件下两代选择育成了籼型温敏型雄性不育株系安农S-1[31].2014年庄楚雄和曹晓风团队发现其温敏特性由tms5位点控制,TMS5编码的RNase ZS1是一种RNA酶,可以特异性地降解泛素-60S核糖体蛋白L40基因(ubiquitin-60S ribosomal protein L40 family, UbL40)的mRNA.在安农S-1中,TMS5基因的一个T到A的单碱基突变导致了该基因在第71个碱基对处提前终止.推测安农S-1中的RNase ZS1功能缺失,致使高温下UbL40的转录本过度积累,最终导致雄性不育[32]. ...
... 虽然农垦58S是大多数光温敏核不育系的亲本来源,但根据周海等[32]的统计,截至2011年,71%的两系杂交组合都依赖于tms5位点,占据了83.8%的两系杂交稻种植面积.张华丽等[53]收集了90种光温敏不育系,通过CAPS标记和RsaⅠ酶切鉴定其携带的光温敏位点,发现衍生自安农S-1和株1S的18个不育系全部携带tms5位点;完全衍生自农垦58S的47个不育系中,只有12个不育系携带了农垦58S的位点pms3,却有29个不育系携带了tms5位点,2个不育系携带了pms3和tms5两个位点,4个不育系既不携带pms3也不携带tms5;其他25个不育系中,17个不育系携带了tms5位点,6个不育系携带了pms3位点,1个不育系携带了pms3和tms5两个位点,1个不育系携带osms1wenmin1位点.这说明虽然很多不育系由农垦58S衍生而来,但它们的不育位点已经从pms3变为了tms5.如广占63S完全由农垦58S衍生而来,但只携带tms5位点而不携带pms3;Y58S的不育位点来自安农S-1和培矮64S,但只携带tms5[53].目前我们尚无法解释遗传谱系和基因位点之间的不一致现象,但推测可能是农垦58S在转育、筛选的过程中混入了含有tms5位点的种子,并被作为优异单株筛选出来所导致的.轮回422S是1990年到1994年间从7001S与轮回422的杂交后代中混收选育得到的温敏不育系,并没有来自安农S-1或株1S的亲缘[54],但轮回422S却携带了其亲本均不具有的tms5位点.以上的研究表明,虽然目前我国光温敏不育系品种众多,但tms5仍是目前两系杂交水稻育种中最依赖的温敏遗传位点. ...
籼型两用不育系衡农S-1育性转换及育性遗传的研究
1
1992
... 1987年,湖南省衡阳市农业科学研究所通过杂交,在长芒野生稻/R0183//测64后代中选育出籼型温敏核不育系衡农S-1[33].1989年该单位将衡农S-1与三系恢复系“明恢63”配组,育成的两系组合“衡两优1号”是早期推广的重要两系品种之一[34].2014年何祖华团队定位到衡农S-1的温敏突变位点[35].2022年,李云海和罗越华团队发现衡农S-1的温敏不育表型受OsMS1wenmin1控制,OsMS1wenmin1与野生型OsMS1均编码组氨酸结合蛋白,可以和转录因子TDR蛋白互作并激活下游基因的表达.野生型OsMS1蛋白定位于细胞核内,但衡农S-1中的一个T到C的单碱基突变改变了OsMS1wenmin1的亚细胞定位与丰度,使其定位于细胞核内和细胞质中.高温下,细胞核内的OsMS1wenmin1水平过低,造成下游基因调控紊乱,最终造成雄性不育[36]. ...
Study on fertility conversion and fertility inheritance of indica TGMS line Hengnong S-1
1
1992
... 1987年,湖南省衡阳市农业科学研究所通过杂交,在长芒野生稻/R0183//测64后代中选育出籼型温敏核不育系衡农S-1[33].1989年该单位将衡农S-1与三系恢复系“明恢63”配组,育成的两系组合“衡两优1号”是早期推广的重要两系品种之一[34].2014年何祖华团队定位到衡农S-1的温敏突变位点[35].2022年,李云海和罗越华团队发现衡农S-1的温敏不育表型受OsMS1wenmin1控制,OsMS1wenmin1与野生型OsMS1均编码组氨酸结合蛋白,可以和转录因子TDR蛋白互作并激活下游基因的表达.野生型OsMS1蛋白定位于细胞核内,但衡农S-1中的一个T到C的单碱基突变改变了OsMS1wenmin1的亚细胞定位与丰度,使其定位于细胞核内和细胞质中.高温下,细胞核内的OsMS1wenmin1水平过低,造成下游基因调控紊乱,最终造成雄性不育[36]. ...
中国两系杂交稻的发展现状及光温敏雄性不育基因研究进展
1
2015
... 1987年,湖南省衡阳市农业科学研究所通过杂交,在长芒野生稻/R0183//测64后代中选育出籼型温敏核不育系衡农S-1[33].1989年该单位将衡农S-1与三系恢复系“明恢63”配组,育成的两系组合“衡两优1号”是早期推广的重要两系品种之一[34].2014年何祖华团队定位到衡农S-1的温敏突变位点[35].2022年,李云海和罗越华团队发现衡农S-1的温敏不育表型受OsMS1wenmin1控制,OsMS1wenmin1与野生型OsMS1均编码组氨酸结合蛋白,可以和转录因子TDR蛋白互作并激活下游基因的表达.野生型OsMS1蛋白定位于细胞核内,但衡农S-1中的一个T到C的单碱基突变改变了OsMS1wenmin1的亚细胞定位与丰度,使其定位于细胞核内和细胞质中.高温下,细胞核内的OsMS1wenmin1水平过低,造成下游基因调控紊乱,最终造成雄性不育[36]. ...
Current situation for development of two-line hybrid rice in China and research progress of rice photoperiod- and thermo- sensitive genic male sterility gene
1
2015
... 1987年,湖南省衡阳市农业科学研究所通过杂交,在长芒野生稻/R0183//测64后代中选育出籼型温敏核不育系衡农S-1[33].1989年该单位将衡农S-1与三系恢复系“明恢63”配组,育成的两系组合“衡两优1号”是早期推广的重要两系品种之一[34].2014年何祖华团队定位到衡农S-1的温敏突变位点[35].2022年,李云海和罗越华团队发现衡农S-1的温敏不育表型受OsMS1wenmin1控制,OsMS1wenmin1与野生型OsMS1均编码组氨酸结合蛋白,可以和转录因子TDR蛋白互作并激活下游基因的表达.野生型OsMS1蛋白定位于细胞核内,但衡农S-1中的一个T到C的单碱基突变改变了OsMS1wenmin1的亚细胞定位与丰度,使其定位于细胞核内和细胞质中.高温下,细胞核内的OsMS1wenmin1水平过低,造成下游基因调控紊乱,最终造成雄性不育[36]. ...
Fine mapping and candidate gene analysis of the novel thermo-sensitive genic male sterility tms9-1 gene in rice
1
2014
... 1987年,湖南省衡阳市农业科学研究所通过杂交,在长芒野生稻/R0183//测64后代中选育出籼型温敏核不育系衡农S-1[33].1989年该单位将衡农S-1与三系恢复系“明恢63”配组,育成的两系组合“衡两优1号”是早期推广的重要两系品种之一[34].2014年何祖华团队定位到衡农S-1的温敏突变位点[35].2022年,李云海和罗越华团队发现衡农S-1的温敏不育表型受OsMS1wenmin1控制,OsMS1wenmin1与野生型OsMS1均编码组氨酸结合蛋白,可以和转录因子TDR蛋白互作并激活下游基因的表达.野生型OsMS1蛋白定位于细胞核内,但衡农S-1中的一个T到C的单碱基突变改变了OsMS1wenmin1的亚细胞定位与丰度,使其定位于细胞核内和细胞质中.高温下,细胞核内的OsMS1wenmin1水平过低,造成下游基因调控紊乱,最终造成雄性不育[36]. ...
A natural allele of OsMS1 responds to temperature changes and confers thermosensitive genic male sterility
2
2022
... 1987年,湖南省衡阳市农业科学研究所通过杂交,在长芒野生稻/R0183//测64后代中选育出籼型温敏核不育系衡农S-1[33].1989年该单位将衡农S-1与三系恢复系“明恢63”配组,育成的两系组合“衡两优1号”是早期推广的重要两系品种之一[34].2014年何祖华团队定位到衡农S-1的温敏突变位点[35].2022年,李云海和罗越华团队发现衡农S-1的温敏不育表型受OsMS1wenmin1控制,OsMS1wenmin1与野生型OsMS1均编码组氨酸结合蛋白,可以和转录因子TDR蛋白互作并激活下游基因的表达.野生型OsMS1蛋白定位于细胞核内,但衡农S-1中的一个T到C的单碱基突变改变了OsMS1wenmin1的亚细胞定位与丰度,使其定位于细胞核内和细胞质中.高温下,细胞核内的OsMS1wenmin1水平过低,造成下游基因调控紊乱,最终造成雄性不育[36]. ...
... 3)构建单碱基突变:通过分析已报道的光温敏不育系可以发现,除了tms5和tms10,大部分光温敏不育位点都是其对应基因的弱等位突变而非敲除突变,例如tms18、tms15和osms1的强等位突变就呈现对温度不敏感的完全雄性不育[36,39,40].目前的证据表明,光温敏不育植株的育性转换温度,很有可能和基因的突变强弱呈现一定的相关性.构建单碱基突变,除了传统的化学诱变等方式,还可以通过基因编辑.目前最前沿的单碱基编辑(base editing)技术,如胞嘧啶编辑器(cytidine base editor, CBE)和腺嘌呤编辑器(adenine base editor, ABE)等,已经实现对单个碱基的精准替换[80-81].韩煜等[40]利用碱基编辑器对TMS15进行单碱基替换,创制了12个光温敏特性不同的不育株系.前人在水稻与拟南芥中的研究表明,许多光温敏不育的表型都是由于某些花粉壁发育所必需的关键功能基因的突变所引起.我们可以对某些具有光温敏基因的重要结构域尝试进行有目的、有选择的单碱基替换,从而获取性状更好、更稳定的光温敏不育株系. ...
Two rice receptor-like kinases maintain male fertility under changing temperatures
2
2017
... 2017年,张大兵团队发现水稻tms10位点.TMS10编码一个亮氨酸受体激酶,与其同源基因TMS10L冗余地调控绒毡层细胞的降解.tms10呈现高温不育、低温可育的表型,而tms10tms10l双突变体呈现不受温度影响的完全雄性不育.在高温下因TMS10突变导致花药绒毡层无法正常降解导致花药外壁缺陷,但低温可以诱导TMS10L大量表达,由此弥补TMS10的功能缺陷并恢复育性[37]. ...
... 张丞等[69]发现,拟南芥中rpg1是一个初生外壁缺陷的温敏不育突变体.低温下因为植物的缓慢发育,使具有冗余功能的同源基因RPG2能够部分恢复rpg1的育性,这与Yu等[37]报道的TMS10L在低温条件下作为冗余基因恢复tms10突变体育性的机制极为相似. ...
Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene
1
2016
... 2022-2023年,杨仲南团队通过EMS诱变和筛选,发现并报道了tms18和tms15位点.OsTMS18编码一个葡萄糖-甲醇-胆碱(GMC)氧化还原酶,因其受到花粉外壁形成关键转录因子OsMS188的直接调控,可能参与了花粉壁的合成.NP1是第三代杂交水稻智能不育系使用的核不育基因[38],tms18是osnp1的弱等位突变体,NP1完全敲除会造成不受温度影响的完全雄性不育,而tms18中一个单碱基突变导致其产生了光温敏表型[39].OsTMS15编码一个富亮氨酸重复序列类受体激酶MSP1,在花药中特异表达,负责启动水稻花药绒毡层的发育.其编码区中TIR区段的一个单碱基突变导致了tms15的温敏表型.tms15的敲除突变体tms15-cr完全缺失绒毡层细胞,tms15中仍然存在绒毡层,但在高温下绒毡层出现空泡化导致不育,而低温下绒毡层发育正常从而恢复植株育性[40].在中花11背景下,ostms18的育性转换温度与tms5相近,而ostms15育性转换温度比tms5更低. ...
Mutation of glucose-methanol-choline oxidoreductase leads to thermosensitive genic male sterility in rice and Arabidopsis
4
2022
... 2022-2023年,杨仲南团队通过EMS诱变和筛选,发现并报道了tms18和tms15位点.OsTMS18编码一个葡萄糖-甲醇-胆碱(GMC)氧化还原酶,因其受到花粉外壁形成关键转录因子OsMS188的直接调控,可能参与了花粉壁的合成.NP1是第三代杂交水稻智能不育系使用的核不育基因[38],tms18是osnp1的弱等位突变体,NP1完全敲除会造成不受温度影响的完全雄性不育,而tms18中一个单碱基突变导致其产生了光温敏表型[39].OsTMS15编码一个富亮氨酸重复序列类受体激酶MSP1,在花药中特异表达,负责启动水稻花药绒毡层的发育.其编码区中TIR区段的一个单碱基突变导致了tms15的温敏表型.tms15的敲除突变体tms15-cr完全缺失绒毡层细胞,tms15中仍然存在绒毡层,但在高温下绒毡层出现空泡化导致不育,而低温下绒毡层发育正常从而恢复植株育性[40].在中花11背景下,ostms18的育性转换温度与tms5相近,而ostms15育性转换温度比tms5更低. ...
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
... 光照和温度是植物生长发育最重要环境因子,低温或短日照条件下植物生长发育速度的减缓是一种普遍现象.水稻在低温或短日照条件下也有发育减缓的现象[57].ostms18表现出温敏不育的表型,其在拟南芥中的同源突变体attms18也存在温敏的表型,具体表现为24℃下可育,28℃下不育.另外,AtTMS18蛋白也与OsTMS18一样定位于绒毡层,这暗示着水稻和拟南芥很有可能具有类似保守的温敏恢复机制[39]. ...
... 3)构建单碱基突变:通过分析已报道的光温敏不育系可以发现,除了tms5和tms10,大部分光温敏不育位点都是其对应基因的弱等位突变而非敲除突变,例如tms18、tms15和osms1的强等位突变就呈现对温度不敏感的完全雄性不育[36,39,40].目前的证据表明,光温敏不育植株的育性转换温度,很有可能和基因的突变强弱呈现一定的相关性.构建单碱基突变,除了传统的化学诱变等方式,还可以通过基因编辑.目前最前沿的单碱基编辑(base editing)技术,如胞嘧啶编辑器(cytidine base editor, CBE)和腺嘌呤编辑器(adenine base editor, ABE)等,已经实现对单个碱基的精准替换[80-81].韩煜等[40]利用碱基编辑器对TMS15进行单碱基替换,创制了12个光温敏特性不同的不育株系.前人在水稻与拟南芥中的研究表明,许多光温敏不育的表型都是由于某些花粉壁发育所必需的关键功能基因的突变所引起.我们可以对某些具有光温敏基因的重要结构域尝试进行有目的、有选择的单碱基替换,从而获取性状更好、更稳定的光温敏不育株系. ...
Low temperature compensates for defective tapetum initiation to restore the fertility of the novel TGMS line ostms15
5
2023
... 2022-2023年,杨仲南团队通过EMS诱变和筛选,发现并报道了tms18和tms15位点.OsTMS18编码一个葡萄糖-甲醇-胆碱(GMC)氧化还原酶,因其受到花粉外壁形成关键转录因子OsMS188的直接调控,可能参与了花粉壁的合成.NP1是第三代杂交水稻智能不育系使用的核不育基因[38],tms18是osnp1的弱等位突变体,NP1完全敲除会造成不受温度影响的完全雄性不育,而tms18中一个单碱基突变导致其产生了光温敏表型[39].OsTMS15编码一个富亮氨酸重复序列类受体激酶MSP1,在花药中特异表达,负责启动水稻花药绒毡层的发育.其编码区中TIR区段的一个单碱基突变导致了tms15的温敏表型.tms15的敲除突变体tms15-cr完全缺失绒毡层细胞,tms15中仍然存在绒毡层,但在高温下绒毡层出现空泡化导致不育,而低温下绒毡层发育正常从而恢复植株育性[40].在中花11背景下,ostms18的育性转换温度与tms5相近,而ostms15育性转换温度比tms5更低. ...
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
... 1)发掘更多尚未应用的光/温敏不育位点:如tms10、tms18、tms15等.在田间实验中,粳稻背景下的tms15表现出了比tms5更加稳定、彻底的高温不育性[40].然而,即使相同位点在不同的遗传背景下都有可能出现不同的光温敏特性和转换温度,因此我们需将具有潜力的光温敏株系与主要品种进行杂交配组,评估其在不同遗传背景下的育性转换温度、高温不育性、异交结实率和自交繁种能力等重要农艺性状. ...
... 3)构建单碱基突变:通过分析已报道的光温敏不育系可以发现,除了tms5和tms10,大部分光温敏不育位点都是其对应基因的弱等位突变而非敲除突变,例如tms18、tms15和osms1的强等位突变就呈现对温度不敏感的完全雄性不育[36,39,40].目前的证据表明,光温敏不育植株的育性转换温度,很有可能和基因的突变强弱呈现一定的相关性.构建单碱基突变,除了传统的化学诱变等方式,还可以通过基因编辑.目前最前沿的单碱基编辑(base editing)技术,如胞嘧啶编辑器(cytidine base editor, CBE)和腺嘌呤编辑器(adenine base editor, ABE)等,已经实现对单个碱基的精准替换[80-81].韩煜等[40]利用碱基编辑器对TMS15进行单碱基替换,创制了12个光温敏特性不同的不育株系.前人在水稻与拟南芥中的研究表明,许多光温敏不育的表型都是由于某些花粉壁发育所必需的关键功能基因的突变所引起.我们可以对某些具有光温敏基因的重要结构域尝试进行有目的、有选择的单碱基替换,从而获取性状更好、更稳定的光温敏不育株系. ...
... [40]利用碱基编辑器对TMS15进行单碱基替换,创制了12个光温敏特性不同的不育株系.前人在水稻与拟南芥中的研究表明,许多光温敏不育的表型都是由于某些花粉壁发育所必需的关键功能基因的突变所引起.我们可以对某些具有光温敏基因的重要结构域尝试进行有目的、有选择的单碱基替换,从而获取性状更好、更稳定的光温敏不育株系. ...
Using bulked extremes and recessive class to map genes for photoperiod-sensitive genic male sterility in rice
1
1994
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Genetic analysis and primary mapping of pms4, a photoperiod-sensitive genic male sterility gene in rice (Oryza sativa)
1
2008
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
水稻温敏显性核不育基因的遗传分析和分子标记定位
1
1999
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Genetic analysis and location of molecular markers for thermo-sensitive dominant genic sterility genes in rice
1
1999
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Tagging and mapping the thermo-sensitive genic male-sterile gene in rice (Oryza sativa L.) with molecular markers
1
1995
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Molecular mapping of a thermosensitive genetic male sterility gene in rice using bulked segregant analysis
1
1997
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Molecular mapping of a rice gene conditioning thermosensitive genic male sterility using AFLP, RFLP and SSR techniques
1
2000
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Development of a genetic marker linked to a new thermo-sensitive male sterile gene in rice(Oryza sativa L.)
1
2004
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Genetic characterization and fine mapping of a novel thermo-sensitive genic male-sterile gene tms6 in rice (Oryza sativa L.)
1
2005
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
Molecular mapping of a new genic male-sterility gene causing chalky endosperm in rice(Oryza sativa L.)
1
1999
... 除上述已报道的数个位点外,水稻尚有其他已发现的光/温敏位点,如pms2[41]、pms4[42]、tms[43]、tms-1[44]、tms-3[45]、tms4[46]、tms6[47,48]、ms-h[49]等,但目前这些基因位点未能成功克隆,作用机理有待研究. ...
株1S温敏核不育基因的发现及超级杂交早稻育种研究
1
2007
... 中国生产应用的两系核不育系,绝大多数(>95%)由农垦58S、安农S-1、株1S等几种早年选育品种衍生而来[24].株1S是杨远柱从抗罗早///科辐红2号/HA79317-7//02428组合的F2群体中发现的,于1997年育成的偏籼型广亲和温敏不育系[50].从亲缘上看,株1S与安农S-1并无直接关系,但株1S及其衍生的不育系均携带有与安农S-1一样的tms5温敏位点[51].严维等通过全基因组多态性分析发现,株1S及其衍生系与安农S-1及其衍生系,在TMS5区域和上下游各1 Mbp范围内存在多个多态性SNP位点和InDel标记,说明二者在该功能区段的来源并不相同[52]. ...
Discovery of a thermo-sensitive genic male sterility gene in Zhu 1S and research on the super hybrid breeding of early rice
1
2007
... 中国生产应用的两系核不育系,绝大多数(>95%)由农垦58S、安农S-1、株1S等几种早年选育品种衍生而来[24].株1S是杨远柱从抗罗早///科辐红2号/HA79317-7//02428组合的F2群体中发现的,于1997年育成的偏籼型广亲和温敏不育系[50].从亲缘上看,株1S与安农S-1并无直接关系,但株1S及其衍生的不育系均携带有与安农S-1一样的tms5温敏位点[51].严维等通过全基因组多态性分析发现,株1S及其衍生系与安农S-1及其衍生系,在TMS5区域和上下游各1 Mbp范围内存在多个多态性SNP位点和InDel标记,说明二者在该功能区段的来源并不相同[52]. ...
Characterization of an RNase Z nonsense mutation identified exclusively in environment-conditioned genic male sterile rice
1
2014
... 中国生产应用的两系核不育系,绝大多数(>95%)由农垦58S、安农S-1、株1S等几种早年选育品种衍生而来[24].株1S是杨远柱从抗罗早///科辐红2号/HA79317-7//02428组合的F2群体中发现的,于1997年育成的偏籼型广亲和温敏不育系[50].从亲缘上看,株1S与安农S-1并无直接关系,但株1S及其衍生的不育系均携带有与安农S-1一样的tms5温敏位点[51].严维等通过全基因组多态性分析发现,株1S及其衍生系与安农S-1及其衍生系,在TMS5区域和上下游各1 Mbp范围内存在多个多态性SNP位点和InDel标记,说明二者在该功能区段的来源并不相同[52]. ...
基于全基因组多态性的安农S-1与株1S亲缘关系分析
1
2019
... 中国生产应用的两系核不育系,绝大多数(>95%)由农垦58S、安农S-1、株1S等几种早年选育品种衍生而来[24].株1S是杨远柱从抗罗早///科辐红2号/HA79317-7//02428组合的F2群体中发现的,于1997年育成的偏籼型广亲和温敏不育系[50].从亲缘上看,株1S与安农S-1并无直接关系,但株1S及其衍生的不育系均携带有与安农S-1一样的tms5温敏位点[51].严维等通过全基因组多态性分析发现,株1S及其衍生系与安农S-1及其衍生系,在TMS5区域和上下游各1 Mbp范围内存在多个多态性SNP位点和InDel标记,说明二者在该功能区段的来源并不相同[52]. ...
Analysis of the genetic relationship between annong S-1 and Zhu 1S based on genome-wide polymmophism
1
2019
... 中国生产应用的两系核不育系,绝大多数(>95%)由农垦58S、安农S-1、株1S等几种早年选育品种衍生而来[24].株1S是杨远柱从抗罗早///科辐红2号/HA79317-7//02428组合的F2群体中发现的,于1997年育成的偏籼型广亲和温敏不育系[50].从亲缘上看,株1S与安农S-1并无直接关系,但株1S及其衍生的不育系均携带有与安农S-1一样的tms5温敏位点[51].严维等通过全基因组多态性分析发现,株1S及其衍生系与安农S-1及其衍生系,在TMS5区域和上下游各1 Mbp范围内存在多个多态性SNP位点和InDel标记,说明二者在该功能区段的来源并不相同[52]. ...
中国两系杂交水稻光温敏核不育基因的鉴定与演化分析
2
2015
... 虽然农垦58S是大多数光温敏核不育系的亲本来源,但根据周海等[32]的统计,截至2011年,71%的两系杂交组合都依赖于tms5位点,占据了83.8%的两系杂交稻种植面积.张华丽等[53]收集了90种光温敏不育系,通过CAPS标记和RsaⅠ酶切鉴定其携带的光温敏位点,发现衍生自安农S-1和株1S的18个不育系全部携带tms5位点;完全衍生自农垦58S的47个不育系中,只有12个不育系携带了农垦58S的位点pms3,却有29个不育系携带了tms5位点,2个不育系携带了pms3和tms5两个位点,4个不育系既不携带pms3也不携带tms5;其他25个不育系中,17个不育系携带了tms5位点,6个不育系携带了pms3位点,1个不育系携带了pms3和tms5两个位点,1个不育系携带osms1wenmin1位点.这说明虽然很多不育系由农垦58S衍生而来,但它们的不育位点已经从pms3变为了tms5.如广占63S完全由农垦58S衍生而来,但只携带tms5位点而不携带pms3;Y58S的不育位点来自安农S-1和培矮64S,但只携带tms5[53].目前我们尚无法解释遗传谱系和基因位点之间的不一致现象,但推测可能是农垦58S在转育、筛选的过程中混入了含有tms5位点的种子,并被作为优异单株筛选出来所导致的.轮回422S是1990年到1994年间从7001S与轮回422的杂交后代中混收选育得到的温敏不育系,并没有来自安农S-1或株1S的亲缘[54],但轮回422S却携带了其亲本均不具有的tms5位点.以上的研究表明,虽然目前我国光温敏不育系品种众多,但tms5仍是目前两系杂交水稻育种中最依赖的温敏遗传位点. ...
... [53].目前我们尚无法解释遗传谱系和基因位点之间的不一致现象,但推测可能是农垦58S在转育、筛选的过程中混入了含有tms5位点的种子,并被作为优异单株筛选出来所导致的.轮回422S是1990年到1994年间从7001S与轮回422的杂交后代中混收选育得到的温敏不育系,并没有来自安农S-1或株1S的亲缘[54],但轮回422S却携带了其亲本均不具有的tms5位点.以上的研究表明,虽然目前我国光温敏不育系品种众多,但tms5仍是目前两系杂交水稻育种中最依赖的温敏遗传位点. ...
Identification and transition analysis of photo-/ thermo-sensitive genic male sterile genes in two-line hybrid rice in China
2
2015
... 虽然农垦58S是大多数光温敏核不育系的亲本来源,但根据周海等[32]的统计,截至2011年,71%的两系杂交组合都依赖于tms5位点,占据了83.8%的两系杂交稻种植面积.张华丽等[53]收集了90种光温敏不育系,通过CAPS标记和RsaⅠ酶切鉴定其携带的光温敏位点,发现衍生自安农S-1和株1S的18个不育系全部携带tms5位点;完全衍生自农垦58S的47个不育系中,只有12个不育系携带了农垦58S的位点pms3,却有29个不育系携带了tms5位点,2个不育系携带了pms3和tms5两个位点,4个不育系既不携带pms3也不携带tms5;其他25个不育系中,17个不育系携带了tms5位点,6个不育系携带了pms3位点,1个不育系携带了pms3和tms5两个位点,1个不育系携带osms1wenmin1位点.这说明虽然很多不育系由农垦58S衍生而来,但它们的不育位点已经从pms3变为了tms5.如广占63S完全由农垦58S衍生而来,但只携带tms5位点而不携带pms3;Y58S的不育位点来自安农S-1和培矮64S,但只携带tms5[53].目前我们尚无法解释遗传谱系和基因位点之间的不一致现象,但推测可能是农垦58S在转育、筛选的过程中混入了含有tms5位点的种子,并被作为优异单株筛选出来所导致的.轮回422S是1990年到1994年间从7001S与轮回422的杂交后代中混收选育得到的温敏不育系,并没有来自安农S-1或株1S的亲缘[54],但轮回422S却携带了其亲本均不具有的tms5位点.以上的研究表明,虽然目前我国光温敏不育系品种众多,但tms5仍是目前两系杂交水稻育种中最依赖的温敏遗传位点. ...
... [53].目前我们尚无法解释遗传谱系和基因位点之间的不一致现象,但推测可能是农垦58S在转育、筛选的过程中混入了含有tms5位点的种子,并被作为优异单株筛选出来所导致的.轮回422S是1990年到1994年间从7001S与轮回422的杂交后代中混收选育得到的温敏不育系,并没有来自安农S-1或株1S的亲缘[54],但轮回422S却携带了其亲本均不具有的tms5位点.以上的研究表明,虽然目前我国光温敏不育系品种众多,但tms5仍是目前两系杂交水稻育种中最依赖的温敏遗传位点. ...
光敏核不育系轮回422S的选育
1
1995
... 虽然农垦58S是大多数光温敏核不育系的亲本来源,但根据周海等[32]的统计,截至2011年,71%的两系杂交组合都依赖于tms5位点,占据了83.8%的两系杂交稻种植面积.张华丽等[53]收集了90种光温敏不育系,通过CAPS标记和RsaⅠ酶切鉴定其携带的光温敏位点,发现衍生自安农S-1和株1S的18个不育系全部携带tms5位点;完全衍生自农垦58S的47个不育系中,只有12个不育系携带了农垦58S的位点pms3,却有29个不育系携带了tms5位点,2个不育系携带了pms3和tms5两个位点,4个不育系既不携带pms3也不携带tms5;其他25个不育系中,17个不育系携带了tms5位点,6个不育系携带了pms3位点,1个不育系携带了pms3和tms5两个位点,1个不育系携带osms1wenmin1位点.这说明虽然很多不育系由农垦58S衍生而来,但它们的不育位点已经从pms3变为了tms5.如广占63S完全由农垦58S衍生而来,但只携带tms5位点而不携带pms3;Y58S的不育位点来自安农S-1和培矮64S,但只携带tms5[53].目前我们尚无法解释遗传谱系和基因位点之间的不一致现象,但推测可能是农垦58S在转育、筛选的过程中混入了含有tms5位点的种子,并被作为优异单株筛选出来所导致的.轮回422S是1990年到1994年间从7001S与轮回422的杂交后代中混收选育得到的温敏不育系,并没有来自安农S-1或株1S的亲缘[54],但轮回422S却携带了其亲本均不具有的tms5位点.以上的研究表明,虽然目前我国光温敏不育系品种众多,但tms5仍是目前两系杂交水稻育种中最依赖的温敏遗传位点. ...
Selection of photoperiod-sensitive male sterile line Lunhui 422S
1
1995
... 虽然农垦58S是大多数光温敏核不育系的亲本来源,但根据周海等[32]的统计,截至2011年,71%的两系杂交组合都依赖于tms5位点,占据了83.8%的两系杂交稻种植面积.张华丽等[53]收集了90种光温敏不育系,通过CAPS标记和RsaⅠ酶切鉴定其携带的光温敏位点,发现衍生自安农S-1和株1S的18个不育系全部携带tms5位点;完全衍生自农垦58S的47个不育系中,只有12个不育系携带了农垦58S的位点pms3,却有29个不育系携带了tms5位点,2个不育系携带了pms3和tms5两个位点,4个不育系既不携带pms3也不携带tms5;其他25个不育系中,17个不育系携带了tms5位点,6个不育系携带了pms3位点,1个不育系携带了pms3和tms5两个位点,1个不育系携带osms1wenmin1位点.这说明虽然很多不育系由农垦58S衍生而来,但它们的不育位点已经从pms3变为了tms5.如广占63S完全由农垦58S衍生而来,但只携带tms5位点而不携带pms3;Y58S的不育位点来自安农S-1和培矮64S,但只携带tms5[53].目前我们尚无法解释遗传谱系和基因位点之间的不一致现象,但推测可能是农垦58S在转育、筛选的过程中混入了含有tms5位点的种子,并被作为优异单株筛选出来所导致的.轮回422S是1990年到1994年间从7001S与轮回422的杂交后代中混收选育得到的温敏不育系,并没有来自安农S-1或株1S的亲缘[54],但轮回422S却携带了其亲本均不具有的tms5位点.以上的研究表明,虽然目前我国光温敏不育系品种众多,但tms5仍是目前两系杂交水稻育种中最依赖的温敏遗传位点. ...
The contributions of sporophytic tapetum to pollen formation
1
2022
... 在花药发育过程中,从小孢子母细胞到花粉形成发生一系列的细胞壁转换过程.一般植物细胞壁主要由纤维素和果胶组成,随着小孢子母细胞的发育,其细胞壁中的纤维素逐渐降解,只留下果胶为主的细胞壁(pectin wall).减数分裂起始前,在果胶壁和质膜之间会形成由β-1,3-葡聚糖组成胼胝质壁(callose wall).种子植物在减数分裂结束后会形成一个四分体(tetrad)结构,四分体壁由果胶壁和胼胝质壁组成.在四分体的中后期,初生外壁(primexine)开始在四分体的下层形成,作为孢粉素沉积的载体.小孢子从四分体释放后,花粉壁开始形成.花粉壁由花粉外壁(exine)和花粉内壁(intine)组成.花粉外壁由孢子体控制,进一步分为外壁外层(sexine)和外壁内层(nexine).花粉外壁外层的主要成分是孢粉素(sporopollenin),具有保护小孢子/花粉基因组防止紫外损伤的作用.花粉内壁由小孢子自身基因组控制合成,与一般植物细胞壁组分相似,主要由纤维素和果胶组成.最后,绒毡层的降解产物会沉积并镶嵌在花粉外壁的缝隙中,形成含油层(tryphine)或称为花粉包被(pollen coat),形成花粉壁.综上所述,随着花药发育,花粉在形成过程中,其细胞壁经过一系列转换过程[55,56]. ...
Pollen wall pattern in Arabidopsis
1
2016
... 在花药发育过程中,从小孢子母细胞到花粉形成发生一系列的细胞壁转换过程.一般植物细胞壁主要由纤维素和果胶组成,随着小孢子母细胞的发育,其细胞壁中的纤维素逐渐降解,只留下果胶为主的细胞壁(pectin wall).减数分裂起始前,在果胶壁和质膜之间会形成由β-1,3-葡聚糖组成胼胝质壁(callose wall).种子植物在减数分裂结束后会形成一个四分体(tetrad)结构,四分体壁由果胶壁和胼胝质壁组成.在四分体的中后期,初生外壁(primexine)开始在四分体的下层形成,作为孢粉素沉积的载体.小孢子从四分体释放后,花粉壁开始形成.花粉壁由花粉外壁(exine)和花粉内壁(intine)组成.花粉外壁由孢子体控制,进一步分为外壁外层(sexine)和外壁内层(nexine).花粉外壁外层的主要成分是孢粉素(sporopollenin),具有保护小孢子/花粉基因组防止紫外损伤的作用.花粉内壁由小孢子自身基因组控制合成,与一般植物细胞壁组分相似,主要由纤维素和果胶组成.最后,绒毡层的降解产物会沉积并镶嵌在花粉外壁的缝隙中,形成含油层(tryphine)或称为花粉包被(pollen coat),形成花粉壁.综上所述,随着花药发育,花粉在形成过程中,其细胞壁经过一系列转换过程[55,56]. ...
Slowing development restores the fertility of thermo-sensitive male-sterile plant lines
4
2020
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
... [57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
... 朱骏等[67,68]以拟南芥温敏不育突变体rvms-1 (reversible male sterile-1)为研究对象,发现低温下的小孢子发育进程被减缓.进一步发现编码细胞周期蛋白激酶的CDKA;1 (A-TYPE CYCLIN- DEPENDENT KINASE;1)点突变后可以恢复rvms-1的高温育性.因此将CDKA;1命名为恢复子1(RESTORER OF RVMS 1, RES1).该基因是雄性配子体发生过程中细胞分裂所必需的.CDKA;1点突变后导致细胞减数分裂用时增加,延长了小孢子的发育时间.该恢复子还可以恢复cals5-6、rpg1、acos5-2等多个温敏突变体在常温下的育性.因此,低温下缓慢发育是温敏不育系育性恢复的共性机制[57].张丞等[69]发现rvms-1、cals5-6、npu、cyp703a2等拟南芥温敏突变体在短光周期或低光照强度的条件下也可以恢复育性.短光照或低光强度的条件与低温情况类似,拟南芥花药发育都明显减缓.因此,这些温敏不育系也是光敏不育系.光照和温度都通过缓慢发育来恢复这些不育系的育性. ...
... 光照和温度是植物生长发育最重要环境因子,低温或短日照条件下植物生长发育速度的减缓是一种普遍现象.水稻在低温或短日照条件下也有发育减缓的现象[57].ostms18表现出温敏不育的表型,其在拟南芥中的同源突变体attms18也存在温敏的表型,具体表现为24℃下可育,28℃下不育.另外,AtTMS18蛋白也与OsTMS18一样定位于绒毡层,这暗示着水稻和拟南芥很有可能具有类似保守的温敏恢复机制[39]. ...
A cellular mechanism underlying the restoration of thermo/photoperiod- sensitive genic male sterility
3
2021
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
... 温敏不育系rvms-2恢复子res2、res3的筛选,进一步解析了缓慢发育恢复光温敏不育系育性的机制.RES2/QRT3编码一种聚半乳糖醛酸酶来分解四分体的果胶壁.在res2 rvms-2中,延迟的果胶壁为小孢子的发育提供了额外的保护,从而恢复rvms-2的育性[58].RES3/UPEX1编码一种阿拉伯半乳糖β-(1,3)-半乳糖基转移酶.在res3中,胼胝质酶A6从绒毡层到药室腔的分泌被滞后,从而延迟了四分体胼胝质壁的降解,这表明延迟降解的四分体胼胝质壁同样保护了rvms-2小孢子的发育,从而恢复育性[70].res2和res3不仅可以恢复rvms-2的育性,也可以恢复其他温敏不育系的育性,说明光温敏不育系通过四分体壁降解的延迟恢复育性是一个普遍机制[58,70].光温敏不育系由于基因的突变,导致自身的细胞壁保护能力降低,而低温下育性得以恢复表明缓慢发育降低了小孢子对细胞壁保护作用的要求(图1). ...
... [58,70].光温敏不育系由于基因的突变,导致自身的细胞壁保护能力降低,而低温下育性得以恢复表明缓慢发育降低了小孢子对细胞壁保护作用的要求(图1). ...
Callose synthase (CalS5) is required for exine formation during microgametogenesis and for pollen viability in Arabidopsis
1
2005
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
No primexine and plasma membrane undulation is essential for primexine deposition and plasma membrane undulation during microsporogenesis in Arabidopsis
1
2012
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
Arabidopsis RPG1 is important for primexine deposition and functions redundantly with RPG2 for plant fertility at the late reproductive stage
1
2013
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
A novel fatty Acyl-CoA Synthetase is required for pollen development and sporopollenin biosynthesis in Arabidopsis
1
2009
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
CYP703 is an ancient cytochrome P450 in land plants catalyzing in-chain hydroxylation of lauric acid to provide building blocks for sporopollenin synthesis in pollen
1
2007
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
New views of tapetum ultrastructure and pollen exine development in Arabidopsis thaliana
1
2014
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
Slowing development facilitates Arabidopsis mgt mutants to accumulate enough magnesium for pollen formation and fertility restoration
1
2020
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
PERSISTENT TAPETAL CELL1 encodes a PHD-finger protein that is required for tapetal cell death and pollen development in rice
1
2011
... 目前拟南芥中已报道了众多的光温敏不育系,如cals5-6、rpg1、npu-2、acos5-2、cyp703a2-1、abcg26-1、rvms-1、mgt5等[57,58].这些基因均直接或间接与细胞壁转换相关,其突变体大多表现出细胞壁转换的缺陷.其中,CALS5负责四分体胼胝质壁的形成[59];RPG1、NPU与初生外壁的形成和质膜波动有关[60,61];脂酰辅酶A合成酶基因ACOS5和细胞色素P450家族基因CYP703A3分别参与孢粉素中长链脂酰辅酶A的合成和长链脂肪酸的羧化[62,63];ABCG26编码一种ATP结合盒转运蛋白,负责孢粉素前体物质转运[64],间接参与了花粉外壁的合成;RVMS参与质膜的形成,间接影响花粉外壁内层的形成[57];MGT5负责镁离子转运调控花粉内壁形成[65].在水稻中也发现了外壁发育相关的光温敏位点,如温敏突变体ostms18花粉外壁缺陷,且TMS18的表达受到花粉外壁的关键转录因子OsMS188的直接调控,因此推测其也参与了花粉外壁的合成[39];OsMS1作为转录因子调控了一系列曾被报道与花粉壁形成相关的基因,对花粉壁形成十分重要[66];水稻ostms15中绒毡层起始缺陷,间接影响了花粉外壁的形成[40](图1). ...
Arabidopsis CDKA;1, a cdc2 homologue, controls proliferation of generative cells in male gametogenesis
1
2006
... 朱骏等[67,68]以拟南芥温敏不育突变体rvms-1 (reversible male sterile-1)为研究对象,发现低温下的小孢子发育进程被减缓.进一步发现编码细胞周期蛋白激酶的CDKA;1 (A-TYPE CYCLIN- DEPENDENT KINASE;1)点突变后可以恢复rvms-1的高温育性.因此将CDKA;1命名为恢复子1(RESTORER OF RVMS 1, RES1).该基因是雄性配子体发生过程中细胞分裂所必需的.CDKA;1点突变后导致细胞减数分裂用时增加,延长了小孢子的发育时间.该恢复子还可以恢复cals5-6、rpg1、acos5-2等多个温敏突变体在常温下的育性.因此,低温下缓慢发育是温敏不育系育性恢复的共性机制[57].张丞等[69]发现rvms-1、cals5-6、npu、cyp703a2等拟南芥温敏突变体在短光周期或低光照强度的条件下也可以恢复育性.短光照或低光强度的条件与低温情况类似,拟南芥花药发育都明显减缓.因此,这些温敏不育系也是光敏不育系.光照和温度都通过缓慢发育来恢复这些不育系的育性. ...
The Cdk1/Cdk2homolog CDKA;1 controls the recombination landscape in Arabidopsis
1
2019
... 朱骏等[67,68]以拟南芥温敏不育突变体rvms-1 (reversible male sterile-1)为研究对象,发现低温下的小孢子发育进程被减缓.进一步发现编码细胞周期蛋白激酶的CDKA;1 (A-TYPE CYCLIN- DEPENDENT KINASE;1)点突变后可以恢复rvms-1的高温育性.因此将CDKA;1命名为恢复子1(RESTORER OF RVMS 1, RES1).该基因是雄性配子体发生过程中细胞分裂所必需的.CDKA;1点突变后导致细胞减数分裂用时增加,延长了小孢子的发育时间.该恢复子还可以恢复cals5-6、rpg1、acos5-2等多个温敏突变体在常温下的育性.因此,低温下缓慢发育是温敏不育系育性恢复的共性机制[57].张丞等[69]发现rvms-1、cals5-6、npu、cyp703a2等拟南芥温敏突变体在短光周期或低光照强度的条件下也可以恢复育性.短光照或低光强度的条件与低温情况类似,拟南芥花药发育都明显减缓.因此,这些温敏不育系也是光敏不育系.光照和温度都通过缓慢发育来恢复这些不育系的育性. ...
Slow development allows redundant genes to restore the fertility of rpg1, a TGMS line in Arabidopsis
2
2022
... 朱骏等[67,68]以拟南芥温敏不育突变体rvms-1 (reversible male sterile-1)为研究对象,发现低温下的小孢子发育进程被减缓.进一步发现编码细胞周期蛋白激酶的CDKA;1 (A-TYPE CYCLIN- DEPENDENT KINASE;1)点突变后可以恢复rvms-1的高温育性.因此将CDKA;1命名为恢复子1(RESTORER OF RVMS 1, RES1).该基因是雄性配子体发生过程中细胞分裂所必需的.CDKA;1点突变后导致细胞减数分裂用时增加,延长了小孢子的发育时间.该恢复子还可以恢复cals5-6、rpg1、acos5-2等多个温敏突变体在常温下的育性.因此,低温下缓慢发育是温敏不育系育性恢复的共性机制[57].张丞等[69]发现rvms-1、cals5-6、npu、cyp703a2等拟南芥温敏突变体在短光周期或低光照强度的条件下也可以恢复育性.短光照或低光强度的条件与低温情况类似,拟南芥花药发育都明显减缓.因此,这些温敏不育系也是光敏不育系.光照和温度都通过缓慢发育来恢复这些不育系的育性. ...
... 张丞等[69]发现,拟南芥中rpg1是一个初生外壁缺陷的温敏不育突变体.低温下因为植物的缓慢发育,使具有冗余功能的同源基因RPG2能够部分恢复rpg1的育性,这与Yu等[37]报道的TMS10L在低温条件下作为冗余基因恢复tms10突变体育性的机制极为相似. ...
Delayed callose degradation restores the fertility of multiple P/TGMS lines in Arabidopsis
2
2022
... 温敏不育系rvms-2恢复子res2、res3的筛选,进一步解析了缓慢发育恢复光温敏不育系育性的机制.RES2/QRT3编码一种聚半乳糖醛酸酶来分解四分体的果胶壁.在res2 rvms-2中,延迟的果胶壁为小孢子的发育提供了额外的保护,从而恢复rvms-2的育性[58].RES3/UPEX1编码一种阿拉伯半乳糖β-(1,3)-半乳糖基转移酶.在res3中,胼胝质酶A6从绒毡层到药室腔的分泌被滞后,从而延迟了四分体胼胝质壁的降解,这表明延迟降解的四分体胼胝质壁同样保护了rvms-2小孢子的发育,从而恢复育性[70].res2和res3不仅可以恢复rvms-2的育性,也可以恢复其他温敏不育系的育性,说明光温敏不育系通过四分体壁降解的延迟恢复育性是一个普遍机制[58,70].光温敏不育系由于基因的突变,导致自身的细胞壁保护能力降低,而低温下育性得以恢复表明缓慢发育降低了小孢子对细胞壁保护作用的要求(图1). ...
... ,70].光温敏不育系由于基因的突变,导致自身的细胞壁保护能力降低,而低温下育性得以恢复表明缓慢发育降低了小孢子对细胞壁保护作用的要求(图1). ...
CYP704B1 is a long-chain fatty acid omega-hydroxylase essential for sporopollenin synthesis in pollen of Arabidopsis
1
2009
... 拟南芥中许多温敏不育系,如acos5、cyp703a3、rvms-1、abcg26,在水稻中的同源基因的突变体osacos12、cyp704b1、rms2、pda1均表现出不受环境因素影响的完全雄性不育[71⇓⇓-74].控制衡农S-1光温敏表型的OsMS1wenmin1在拟南芥的同源突变体atms1也未发现温敏现象[75,76].这种差异很可能与花粉壁的缺陷程度以及所处的生长环境恶劣程度有关.鉴于水稻与拟南芥在生长周期、生长环境和物种分类上存在显著差异,水稻可能具有除缓慢发育以外的其他温度敏感机制(图1). ...
OsACOS12, an orthologue of Arabidopsis acyl-CoA synthetase5, plays an important role in pollen exine formation and anther development in rice
1
2016
... 拟南芥中许多温敏不育系,如acos5、cyp703a3、rvms-1、abcg26,在水稻中的同源基因的突变体osacos12、cyp704b1、rms2、pda1均表现出不受环境因素影响的完全雄性不育[71⇓⇓-74].控制衡农S-1光温敏表型的OsMS1wenmin1在拟南芥的同源突变体atms1也未发现温敏现象[75,76].这种差异很可能与花粉壁的缺陷程度以及所处的生长环境恶劣程度有关.鉴于水稻与拟南芥在生长周期、生长环境和物种分类上存在显著差异,水稻可能具有除缓慢发育以外的其他温度敏感机制(图1). ...
RMS2 encoding a GDSL lipase mediates lipid homeostasis in anthers to determine rice male fertility
1
2020
... 拟南芥中许多温敏不育系,如acos5、cyp703a3、rvms-1、abcg26,在水稻中的同源基因的突变体osacos12、cyp704b1、rms2、pda1均表现出不受环境因素影响的完全雄性不育[71⇓⇓-74].控制衡农S-1光温敏表型的OsMS1wenmin1在拟南芥的同源突变体atms1也未发现温敏现象[75,76].这种差异很可能与花粉壁的缺陷程度以及所处的生长环境恶劣程度有关.鉴于水稻与拟南芥在生长周期、生长环境和物种分类上存在显著差异,水稻可能具有除缓慢发育以外的其他温度敏感机制(图1). ...
Post-meiotic deficient anther1 (PDA1) encodes an ABC transporter required for the development of anther cuticle and pollen exine in rice
1
2013
... 拟南芥中许多温敏不育系,如acos5、cyp703a3、rvms-1、abcg26,在水稻中的同源基因的突变体osacos12、cyp704b1、rms2、pda1均表现出不受环境因素影响的完全雄性不育[71⇓⇓-74].控制衡农S-1光温敏表型的OsMS1wenmin1在拟南芥的同源突变体atms1也未发现温敏现象[75,76].这种差异很可能与花粉壁的缺陷程度以及所处的生长环境恶劣程度有关.鉴于水稻与拟南芥在生长周期、生长环境和物种分类上存在显著差异,水稻可能具有除缓慢发育以外的其他温度敏感机制(图1). ...
The Arabidopsis MALE STERILITY1 (MS1) gene is a transcriptional regulator of male gametogenesis, with homology to the PHD-finger family of transcription factors
1
2001
... 拟南芥中许多温敏不育系,如acos5、cyp703a3、rvms-1、abcg26,在水稻中的同源基因的突变体osacos12、cyp704b1、rms2、pda1均表现出不受环境因素影响的完全雄性不育[71⇓⇓-74].控制衡农S-1光温敏表型的OsMS1wenmin1在拟南芥的同源突变体atms1也未发现温敏现象[75,76].这种差异很可能与花粉壁的缺陷程度以及所处的生长环境恶劣程度有关.鉴于水稻与拟南芥在生长周期、生长环境和物种分类上存在显著差异,水稻可能具有除缓慢发育以外的其他温度敏感机制(图1). ...
The MALE STERILITY1gene of Arabidopsis, encoding a nuclear protein with a PHD-finger motif, is expressed in tapetal cells and is required for pollen maturation
1
2002
... 拟南芥中许多温敏不育系,如acos5、cyp703a3、rvms-1、abcg26,在水稻中的同源基因的突变体osacos12、cyp704b1、rms2、pda1均表现出不受环境因素影响的完全雄性不育[71⇓⇓-74].控制衡农S-1光温敏表型的OsMS1wenmin1在拟南芥的同源突变体atms1也未发现温敏现象[75,76].这种差异很可能与花粉壁的缺陷程度以及所处的生长环境恶劣程度有关.鉴于水稻与拟南芥在生长周期、生长环境和物种分类上存在显著差异,水稻可能具有除缓慢发育以外的其他温度敏感机制(图1). ...
水稻温敏核不育系株1S及其衍生系育性对低温的敏感性差异比较研究
1
2021
... 从1973年光温敏现象被发现以来,经过几十年的发展,人们对光温敏现象的理解正在逐步深入.我国的光温敏不育系审定品种数量不断增长,但这些不育系的基因来源却十分单一,绝大多数都受到pms3或tms5位点的控制.随着两系育种的不断发展,tms5逐渐成为了最主流的光温敏位点.两系法制种极其依赖天气,因此需要保证十分彻底的高温不育性和尽可能低的育性转换温度.2000年华中农业大学的人工气候箱育性鉴定结果表明tms5控制的广占63S在高温、长日照下的不育性比pms3控制的培矮64S更好[23],吴亚先等[77]研究了12份温敏不育株系的低温敏感性,发现隆科638S、晶4155S的不育起点温度低于或接近23℃,Y58S的不育起点温度达到了23.0~23.5℃,广占63-4S则高于23.5℃.这也是2005年以后培矮64S逐渐被广占63S取代,广占63S又逐渐被Y58S、隆科638S、晶4155S等品种相继取代,成为应用最广泛的不育系的重要原因之一. ...
Comparative studies on the difference of fertility sensitivity to low temperarure of Zhu 1S and its derived TGMS lines in rice
1
2021
... 从1973年光温敏现象被发现以来,经过几十年的发展,人们对光温敏现象的理解正在逐步深入.我国的光温敏不育系审定品种数量不断增长,但这些不育系的基因来源却十分单一,绝大多数都受到pms3或tms5位点的控制.随着两系育种的不断发展,tms5逐渐成为了最主流的光温敏位点.两系法制种极其依赖天气,因此需要保证十分彻底的高温不育性和尽可能低的育性转换温度.2000年华中农业大学的人工气候箱育性鉴定结果表明tms5控制的广占63S在高温、长日照下的不育性比pms3控制的培矮64S更好[23],吴亚先等[77]研究了12份温敏不育株系的低温敏感性,发现隆科638S、晶4155S的不育起点温度低于或接近23℃,Y58S的不育起点温度达到了23.0~23.5℃,广占63-4S则高于23.5℃.这也是2005年以后培矮64S逐渐被广占63S取代,广占63S又逐渐被Y58S、隆科638S、晶4155S等品种相继取代,成为应用最广泛的不育系的重要原因之一. ...
两系法杂交水稻研究和应用中若干问题的思与行
1
2010
... 但遗憾的是,近二十年来,我国大面积应用的光/温敏不育系仍然高度依赖tms5位点.尽管tms5拥有比pms3更优良的不育性状,但过于单一的遗传位点很可能增加制种的风险.近年来受到各类极端天气的影响,夏季常见的台风、突发的异常低温都可能会对两系制种安全造成严重的威胁,如2009年江苏、安徽、四川等地出现多日24℃左右的低温致使育性波动导致生产事故,造成了严重的经济损失[78].因此,发现和利用不育性更稳定的光温敏核不育系是两系育种的关键.目前,育种家的主流解决方案是以tms5作为基础温敏位点,利用自然和人工环境,加压筛选出育性转换温度较低的不育株系.除此之外,笔者认为还可以采取以下策略: ...
Thoughts and practice on some problems about research and application of two-line hybrid rice
1
2011
... 但遗憾的是,近二十年来,我国大面积应用的光/温敏不育系仍然高度依赖tms5位点.尽管tms5拥有比pms3更优良的不育性状,但过于单一的遗传位点很可能增加制种的风险.近年来受到各类极端天气的影响,夏季常见的台风、突发的异常低温都可能会对两系制种安全造成严重的威胁,如2009年江苏、安徽、四川等地出现多日24℃左右的低温致使育性波动导致生产事故,造成了严重的经济损失[78].因此,发现和利用不育性更稳定的光温敏核不育系是两系育种的关键.目前,育种家的主流解决方案是以tms5作为基础温敏位点,利用自然和人工环境,加压筛选出育性转换温度较低的不育株系.除此之外,笔者认为还可以采取以下策略: ...
水稻光温敏核不育基因tms5与pms3的互作效应
1
2020
... 2)聚合多个位点:王芳权等[79]发现携带pms3与tms5双位点的不育株系,其育性转换温度比只携带tms5位点的株系更低.这提示我们可以在现有光温敏株系的基础上聚合多个光温敏不育位点,以创制更加稳定的光温敏不育系. ...
Interactive effects of the photoperiod-/ thermo-sensitive genic male sterile genes tms5 and pms3 in rice
1
2020
... 2)聚合多个位点:王芳权等[79]发现携带pms3与tms5双位点的不育株系,其育性转换温度比只携带tms5位点的株系更低.这提示我们可以在现有光温敏株系的基础上聚合多个光温敏不育位点,以创制更加稳定的光温敏不育系. ...
Efficient generation of homozygous substitutions in rice in one generation utilizing an rABE8e base editor
1
2021
... 3)构建单碱基突变:通过分析已报道的光温敏不育系可以发现,除了tms5和tms10,大部分光温敏不育位点都是其对应基因的弱等位突变而非敲除突变,例如tms18、tms15和osms1的强等位突变就呈现对温度不敏感的完全雄性不育[36,39,40].目前的证据表明,光温敏不育植株的育性转换温度,很有可能和基因的突变强弱呈现一定的相关性.构建单碱基突变,除了传统的化学诱变等方式,还可以通过基因编辑.目前最前沿的单碱基编辑(base editing)技术,如胞嘧啶编辑器(cytidine base editor, CBE)和腺嘌呤编辑器(adenine base editor, ABE)等,已经实现对单个碱基的精准替换[80-81].韩煜等[40]利用碱基编辑器对TMS15进行单碱基替换,创制了12个光温敏特性不同的不育株系.前人在水稻与拟南芥中的研究表明,许多光温敏不育的表型都是由于某些花粉壁发育所必需的关键功能基因的突变所引起.我们可以对某些具有光温敏基因的重要结构域尝试进行有目的、有选择的单碱基替换,从而获取性状更好、更稳定的光温敏不育株系. ...
Genome editing for plant research and crop improvement
1
2021
... 3)构建单碱基突变:通过分析已报道的光温敏不育系可以发现,除了tms5和tms10,大部分光温敏不育位点都是其对应基因的弱等位突变而非敲除突变,例如tms18、tms15和osms1的强等位突变就呈现对温度不敏感的完全雄性不育[36,39,40].目前的证据表明,光温敏不育植株的育性转换温度,很有可能和基因的突变强弱呈现一定的相关性.构建单碱基突变,除了传统的化学诱变等方式,还可以通过基因编辑.目前最前沿的单碱基编辑(base editing)技术,如胞嘧啶编辑器(cytidine base editor, CBE)和腺嘌呤编辑器(adenine base editor, ABE)等,已经实现对单个碱基的精准替换[80-81].韩煜等[40]利用碱基编辑器对TMS15进行单碱基替换,创制了12个光温敏特性不同的不育株系.前人在水稻与拟南芥中的研究表明,许多光温敏不育的表型都是由于某些花粉壁发育所必需的关键功能基因的突变所引起.我们可以对某些具有光温敏基因的重要结构域尝试进行有目的、有选择的单碱基替换,从而获取性状更好、更稳定的光温敏不育株系. ...
HMS1 interacts with HMS1I to regulate very-long-chain fatty acid biosynthesis and the humidity-sensitive genic male sterility in rice (Oryza sativa)
1
2020
... 4)挖掘新的环境敏感型雄性不育系,如湿度敏感型雄性不育系(humidity-sensitive genic male sterile, HGMS):漆小泉和周海团队分别报道了湿度影响花粉水合从而改变水稻育性的现象[82,83].在一些气候干燥的内陆地区,湿度是相比光照和温度是更加稳定可控的环境因素,湿敏不育系的发现为一些干旱地区农作物的两系制种提供了新的思路. ...
Deficiency of a triterpene pathway results in humidity-sensitive genic male sterility in rice
1
2018
... 4)挖掘新的环境敏感型雄性不育系,如湿度敏感型雄性不育系(humidity-sensitive genic male sterile, HGMS):漆小泉和周海团队分别报道了湿度影响花粉水合从而改变水稻育性的现象[82,83].在一些气候干燥的内陆地区,湿度是相比光照和温度是更加稳定可控的环境因素,湿敏不育系的发现为一些干旱地区农作物的两系制种提供了新的思路. ...


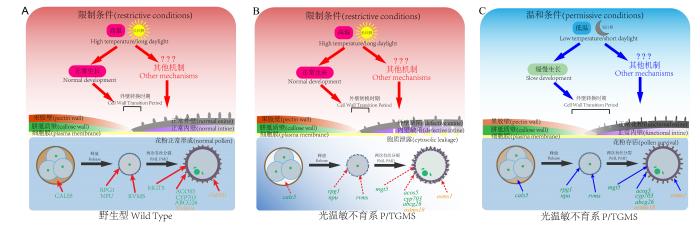

{kind=link}
{kind=link}