Influence of soil type and genotype on Cd bioavailability and uptake by rice and implications for food safety
1
2012
... 植物在生长发育过程中,除需要一定的光照、水分、空气和热量外,还必须不断地从外界吸收所必需的各种营养元素并进行同化,以维持其正常的生命活动.植物通过各种类型的离子转运通道,利用根部从外界吸收并转运这些必需矿质元素,维持正常的生长发育.与此同时,有一些非必需矿质元素也会利用这些通道进入植物体内,如镉(Cd)、铅(Pb)、砷(As)等,这些元素不参与构成植物体的生物结构,也不参与植物体的正常代谢活动,不仅会对植物本身造成伤害,还能通过食物链进入人体,诱发人体疾病,影响人体健康.水稻是世界上重要的粮食作物之一,在我国农业生产中占据着重要地位,超过60%的人口以大米为主食,我国每年水稻的消耗量占谷物消耗量的55%[1].与玉米、小麦和大豆等作物相比,水稻极易富集Cd[2-3],并且耕地酸化或污染可导致土壤中活性Cd含量增加,这又加剧了水稻中Cd的积累,使稻米Cd超标事件频繁发生. ...
Selection and breeding of plant cultivars to minimize cadmium accumulation
1
2008
... 植物在生长发育过程中,除需要一定的光照、水分、空气和热量外,还必须不断地从外界吸收所必需的各种营养元素并进行同化,以维持其正常的生命活动.植物通过各种类型的离子转运通道,利用根部从外界吸收并转运这些必需矿质元素,维持正常的生长发育.与此同时,有一些非必需矿质元素也会利用这些通道进入植物体内,如镉(Cd)、铅(Pb)、砷(As)等,这些元素不参与构成植物体的生物结构,也不参与植物体的正常代谢活动,不仅会对植物本身造成伤害,还能通过食物链进入人体,诱发人体疾病,影响人体健康.水稻是世界上重要的粮食作物之一,在我国农业生产中占据着重要地位,超过60%的人口以大米为主食,我国每年水稻的消耗量占谷物消耗量的55%[1].与玉米、小麦和大豆等作物相比,水稻极易富集Cd[2-3],并且耕地酸化或污染可导致土壤中活性Cd含量增加,这又加剧了水稻中Cd的积累,使稻米Cd超标事件频繁发生. ...
Nramp5 expression and functionality likely explain higher cadmium uptake in rice than in wheat and maize
1
2018
... 植物在生长发育过程中,除需要一定的光照、水分、空气和热量外,还必须不断地从外界吸收所必需的各种营养元素并进行同化,以维持其正常的生命活动.植物通过各种类型的离子转运通道,利用根部从外界吸收并转运这些必需矿质元素,维持正常的生长发育.与此同时,有一些非必需矿质元素也会利用这些通道进入植物体内,如镉(Cd)、铅(Pb)、砷(As)等,这些元素不参与构成植物体的生物结构,也不参与植物体的正常代谢活动,不仅会对植物本身造成伤害,还能通过食物链进入人体,诱发人体疾病,影响人体健康.水稻是世界上重要的粮食作物之一,在我国农业生产中占据着重要地位,超过60%的人口以大米为主食,我国每年水稻的消耗量占谷物消耗量的55%[1].与玉米、小麦和大豆等作物相比,水稻极易富集Cd[2-3],并且耕地酸化或污染可导致土壤中活性Cd含量增加,这又加剧了水稻中Cd的积累,使稻米Cd超标事件频繁发生. ...
Current status of cadmium as an environmental health problem
1
2009
... 人类摄入过量的Cd会严重危害身体健康[4],长期接触Cd会导致肾功能障碍和骨质疏松症等疾病[5-6].例如,日本的“痛痛病”事件,即属最严重的慢性Cd中毒事件.在我国南方部分地区,稻米Cd超标已然成为引起社会广泛关注的食品安全问题.如何减少Cd在水稻籽粒中的积累已经成为水稻生产中亟待解决的关键问题. ...
Mechanisms of cadmium- mediated acute hepatotoxicity
1
2000
... 人类摄入过量的Cd会严重危害身体健康[4],长期接触Cd会导致肾功能障碍和骨质疏松症等疾病[5-6].例如,日本的“痛痛病”事件,即属最严重的慢性Cd中毒事件.在我国南方部分地区,稻米Cd超标已然成为引起社会广泛关注的食品安全问题.如何减少Cd在水稻籽粒中的积累已经成为水稻生产中亟待解决的关键问题. ...
Non-renal effects and the risk assessment of environmental cadmium exposure
1
2014
... 人类摄入过量的Cd会严重危害身体健康[4],长期接触Cd会导致肾功能障碍和骨质疏松症等疾病[5-6].例如,日本的“痛痛病”事件,即属最严重的慢性Cd中毒事件.在我国南方部分地区,稻米Cd超标已然成为引起社会广泛关注的食品安全问题.如何减少Cd在水稻籽粒中的积累已经成为水稻生产中亟待解决的关键问题. ...
Cadmium in rice: Transport mechanisms, influencing factors, and minimizing measures
1
2017
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Glutamate alleviates cadmium toxicity in rice via suppressing cadmium uptake and translocation
1
2020
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Cadmium accumulation in rice straws and derived biochars as affected by metal exposure, soil types and rice genotypes
1
2021
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Effect of cultivation practices on cadmium concentration in rice grain
1
2008
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Effect of land use pattern change from paddy soil to vegetable soil on the adsorption-desorption of cadmium by soil aggregates
1
2017
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Bioaccumulation of cadmium in soil organisms: With focus on wood ash application
1
2018
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Limestone dosage response of cadmium phytoavailability minimization in rice: A trade-off relationship between soil pH and amorphous manganese content
1
2021
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Isolation and characterization of a rice line with high Cd accumulation for potential use in phytoremediation
1
2017
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Phytoremediation of mine tailings with Atriplex halimus and organic/inorganic amendments: A five-year field case study
1
2018
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
水稻镉、铅、砷低含量基因型的筛选
1
2012
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
水稻镉、铅、砷低含量基因型的筛选
1
2012
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
适于轻度Cd、As污染土壤种植的水稻品种筛选
1
2012
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
适于轻度Cd、As污染土壤种植的水稻品种筛选
1
2012
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Preliminary study on rice varieties with low cadmium accumulation by mixed planting
1
2018
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Effective reduction of cadmium accumulation in rice grain by expressing OsHMA3 under the control of the OsHMA2 promoter
1
2018
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
Mutation at different sites of metal transporter gene OsNramp5 affects Cd accumulation and related agronomic traits in rice (Oryza sativa L.)
8
2019
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
20]
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
20]
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
... 随着消费水平的提高,人们对稻米品质的要求也越来越高,安全、健康营养的稻米已成为最基本的需求.Cd富集在水稻中会影响稻米的品质,经人体摄入后会影响健康.有研究表明OsNramp5变异几乎对米质没什么影响,在该研究中突变体的垩白粒率和垩白度小幅下降,蛋白含量轻微增加,而且在一些品种中直链淀粉含量小幅下降,几乎不影响稻米的食味品质[49].Wang等[20]研究表明OsNramp5突变对籽粒中糙米率、精米率、直链淀粉含量、胶稠度和蛋白质含量没有显著影响.随着OsNramp5突变程度的增加,整精米率显著降低,垩白度和垩白粒率显著增加.通过改变OsNramp5的突变位点可以来缓解垩白度和垩白粒率等品质性状.董家瑜等[58]研究表明在正常Mn浓度的大田条件下,以华占为背景的osnramp5突变体的稻米品质较野生型均无显著差异,但以中花11为背景的突变体的胶稠度降低;且开花期高温造成以中花11为背景的的osnramp5突变体的垩白粒率和垩白度上升;在低Mn浓度大田条件下,2种遗传背景的osnramp5突变体的垩白粒率和垩白度上升.在水稻中,OsNramp5同时影响Cd和Mn的吸收和转运,OsNramp5变异降低水稻植株中Cd积累量的同时,也会降低Mn的含量.研究结果显示,高Mn环境可以一定程度改善由OsNramp5变异导致的米质变劣问题[58].因此,在后续利用OsNramp5基因变异选育Cd低积累水稻品种及实际生产应用中,应当将土壤Mn含量纳入品种适用性的考虑范围. ...
Overexpression of the manganese/cadmium transporter OsNRAMP5 reduces cadmium accumulation in rice grain
3
2020
... 目前,水稻Cd污染的控制途径主要有四种:一是通过优化栽培管理措施来抑制土壤中可交换Cd的形成[7⇓-9];二是通过化学或者生物学方法,改变土壤中Cd的现有形态或去除土壤中的Cd,从而阻碍稻田土壤中的Cd通过根系进入植物体内[10⇓⇓-13];三是利用高积累作物将Cd转移到地上部,从而减少土壤中Cd的含量[14-15];四是培育Cd低积累水稻品种[16⇓⇓⇓⇓-21].其中,培育Cd低积累水稻品种是解决稻米Cd超标问题最直接和经济的方法. ...
... 值得一提的是,多数研究发现敲除OsNramp5后能降低水稻中的Cd含量,而Chang等[21]在BG367和中花11的背景下,利用OsActin1和玉米Ubiquitin启动子驱动OsNramp5基因,发现过表达株系在根尖和侧根原基积累了更多的Cd,但减少了其分配到地上部的比例,显著降低了地上部的Cd含量,籽粒中Cd含量比野生型降低了49%~94%,推测OsNramp5过表达植株是由于Cd向中柱的径向运输被破坏,从而减少了Cd从根到地上部的转运. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
The NRAMP family of metal-ion transporters
1
2006
... 天然抗性相关巨噬细胞蛋白(NRAMP)家族成员作为膜蛋白与金属离子转运密切相关.NRAMP家族基因在包括细菌、酵母、藻类、植物和动物在内的所有生物体进化中都比较保守[22],大约具有25%~35%的序列同源性[23].这种保守性说明选择压力使生物体中NRAMP家族基因实现了相同的功能,即跨细胞膜运输金属离子.NRAMP蛋白家族作为一种二价金属转运体,参与多种金属离子的运输,如铁(Fe2+)、锰(Mn2+)和镉(Cd2+)离子[24-25]. ...
Molecular mechanism of NRAMP-family transition metal transport
1
2021
... 天然抗性相关巨噬细胞蛋白(NRAMP)家族成员作为膜蛋白与金属离子转运密切相关.NRAMP家族基因在包括细菌、酵母、藻类、植物和动物在内的所有生物体进化中都比较保守[22],大约具有25%~35%的序列同源性[23].这种保守性说明选择压力使生物体中NRAMP家族基因实现了相同的功能,即跨细胞膜运输金属离子.NRAMP蛋白家族作为一种二价金属转运体,参与多种金属离子的运输,如铁(Fe2+)、锰(Mn2+)和镉(Cd2+)离子[24-25]. ...
Genome-wide analysis of eukaryote thaumatin-like proteins (TLPs) with an emphasis on poplar
1
2011
... 天然抗性相关巨噬细胞蛋白(NRAMP)家族成员作为膜蛋白与金属离子转运密切相关.NRAMP家族基因在包括细菌、酵母、藻类、植物和动物在内的所有生物体进化中都比较保守[22],大约具有25%~35%的序列同源性[23].这种保守性说明选择压力使生物体中NRAMP家族基因实现了相同的功能,即跨细胞膜运输金属离子.NRAMP蛋白家族作为一种二价金属转运体,参与多种金属离子的运输,如铁(Fe2+)、锰(Mn2+)和镉(Cd2+)离子[24-25]. ...
Metal ion transporters and homeostasis
1
1999
... 天然抗性相关巨噬细胞蛋白(NRAMP)家族成员作为膜蛋白与金属离子转运密切相关.NRAMP家族基因在包括细菌、酵母、藻类、植物和动物在内的所有生物体进化中都比较保守[22],大约具有25%~35%的序列同源性[23].这种保守性说明选择压力使生物体中NRAMP家族基因实现了相同的功能,即跨细胞膜运输金属离子.NRAMP蛋白家族作为一种二价金属转运体,参与多种金属离子的运输,如铁(Fe2+)、锰(Mn2+)和镉(Cd2+)离子[24-25]. ...
Cloning of three ZIP/Nramp transporter genes from a Ni hyperaccumulator plant Thlaspi japonicum and their Ni2+-transport abilities
1
2005
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
Functional characterization of NRAMP3 and NRAMP4 from the metal hyperaccumulator Thlaspi caerulescens
1
2009
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
The Thlaspi caerulescens NRAMP homologue TcNRAMP3 is capable of divalent cation transport
1
2009
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
The iron-regulated transporter, MbNRAMP1, isolated from Malus baccata is involved in Fe, Mn and Cd trafficking
1
2008
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
A major quantitative trait locus for increasing cadmium-specific concentration in rice grain is located on the short arm of chromosome 7
2
2010
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
The OsNRAMP1 iron transporter is involved in Cd accumulation in rice
2
2011
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
The tonoplast-localized transporter OsNRAMP2 is involved in iron homeostasis and affects seed germination in rice
2
2021
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
A node-based switch for preferential distribution of manganese in rice
2
2013
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
OsNRAMP3 is a vascular bundles-specific manganese transporter that is responsible for manganese distribution in rice
2
2013
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
Plasma membrane-localized transporter for aluminum in rice
2
2010
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
Natural variation underlies alterations in Nramp aluminum transporter (NRAT1) expression and function that play a key role in rice aluminum tolerance
2
2014
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
Two NRAMP6 isoforms function as iron and manganese transporters and contribute to disease resistance in rice
2
2017
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
水稻OsNRAMP7基因的克隆、表达及生物信息学分析
2
2021
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
水稻OsNRAMP7基因的克隆、表达及生物信息学分析
2
2021
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
Identification of putative target genes to manipulate Fe and Zn concentrations in rice grains
2
2010
... 科学家已在多种植物中鉴定到NRAMP家族基因[26⇓⇓-29].在水稻中共鉴定到7个,分别为OsNramp1、OsNramp2、OsNramp3、OsNramp4、OsNramp5、OsNramp6和OsNramp7(表1).其中,OsNramp1在质膜上充当Cd内流转运体,在Cd从根到茎的转运过程中起关键作用,主要在水稻根系中表达,且不受Cd诱导,但在缺Fe条件下表达高度上调[30-31].OsNramp2定位于液泡膜,参与水稻Fe稳态,将Fe从液泡运输到细胞质中,在种子萌发过程中起着关键作用[32].OsNramp3编码质膜定位蛋白,在维管束特别是韧皮部细胞中特异表达,是一种响应环境中Mn含量变化的Mn转运调节器,并参与了Mn从老叶到幼叶的再分配[33,34].OsNramp4编码一个定位于根细胞质膜的三价铝(Al)转运蛋白,参与Al3+的转运[35-36].OsNramp6定位于质膜,具有Fe和Mn转运蛋白的功能,且与水稻生长及抗病性相关[37].而OsNramp7可能与非生物胁迫及Fe、Zn转运相关[38-39]. ...
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice
7
2012
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... 日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... [40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice
8
2012
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... 日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
41]
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
41]
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
OsNRAMP5, a major player for constitutive iron and manganese uptake in rice
4
2012
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... 日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... [42-43,46 -47,55]. ...
OsNRAMP5 contributes to manganese translocation and distribution in rice shoots
6
2014
... NRAMP protein family genes in rice.
Table 1. 基因符号
Gene symbol | 组织表达
Tissue expression | 亚细胞定位
Subcellular localization | 金属转运功能
Metal transport function | 参考文献
Reference |
| OsNramp1 | 根、叶 Root, leaf | 质膜Plasma membrane | Cd | [30,31] |
| OsNramp2 | 地上部 Aboveground part | 液泡膜 Tonoplast | Fe | [32] |
| OsNramp3 | 维管束 Vascular bundle | 质膜Plasma membrane | Mn | [33,34] |
| OsNramp4 | 根 Root | 质膜Plasma membrane | Al | [35,36] |
| OsNramp5 | 根 Root | 质膜Plasma membrane | Cd, Mn, Fe | [40⇓⇓-43] |
| OsNramp6 | 不详 Unavailable | 质膜Plasma membrane | Fe, Mn | [37] |
| OsNramp7 | 根、茎、幼穗 Root, culm, young panicle | 不详 Unavailable | Fe, Zn | [38,39] |
日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... 日本冈山大学马建锋团队从水稻品种中花11中克隆了OSNRAMP5的全长cDNA[40],该基因具有13个外显子和12个内含子,编码区全长6874 bp,其中CDS全长1617 bp(图1).OsNRAMP5编码蛋白定位在质膜上,分布于根外皮层和根内皮层的远端,是水稻根部细胞参与吸收二价金属阳离子的主要转运蛋白,参与外源Cd2+、Mn2+和Fe2+的吸收,也负责这些离子从根向地上部的运输[41⇓-43].OsNramp5是近年来水稻中发现的唯一一个具有高亲和能力的Cd吸收转运子,该基因的突变能极大地限制水稻对Cd的吸收[40].近年来,利用该基因突变阻断水稻根系细胞对土壤Cd的吸收来降低水稻Cd含量引起水稻遗传育种家的关注(表2). ...
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... -43,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
Cadmium transport and tolerance in rice: Perspectives for reducing grain cadmium accumulation
1
2012
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
1
2014
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
1
2014
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield
6
2017
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
46]
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... ,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
Reduction in cadmium accumulation in japonica rice grains by CRISPR/Cas9- mediated editing of OsNRAMP5
7
2019
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
47]
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
47]
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
Characterization and evaluation of OsLCT1 and OsNramp5 mutants generated through CRISPR/Cas9-mediated mutagenesis for breeding low Cd rice
5
2019
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
48]
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
利用CRISPR/Cas9敲除OsNramp5基因创制低镉籼稻
5
2019
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
... 随着消费水平的提高,人们对稻米品质的要求也越来越高,安全、健康营养的稻米已成为最基本的需求.Cd富集在水稻中会影响稻米的品质,经人体摄入后会影响健康.有研究表明OsNramp5变异几乎对米质没什么影响,在该研究中突变体的垩白粒率和垩白度小幅下降,蛋白含量轻微增加,而且在一些品种中直链淀粉含量小幅下降,几乎不影响稻米的食味品质[49].Wang等[20]研究表明OsNramp5突变对籽粒中糙米率、精米率、直链淀粉含量、胶稠度和蛋白质含量没有显著影响.随着OsNramp5突变程度的增加,整精米率显著降低,垩白度和垩白粒率显著增加.通过改变OsNramp5的突变位点可以来缓解垩白度和垩白粒率等品质性状.董家瑜等[58]研究表明在正常Mn浓度的大田条件下,以华占为背景的osnramp5突变体的稻米品质较野生型均无显著差异,但以中花11为背景的突变体的胶稠度降低;且开花期高温造成以中花11为背景的的osnramp5突变体的垩白粒率和垩白度上升;在低Mn浓度大田条件下,2种遗传背景的osnramp5突变体的垩白粒率和垩白度上升.在水稻中,OsNramp5同时影响Cd和Mn的吸收和转运,OsNramp5变异降低水稻植株中Cd积累量的同时,也会降低Mn的含量.研究结果显示,高Mn环境可以一定程度改善由OsNramp5变异导致的米质变劣问题[58].因此,在后续利用OsNramp5基因变异选育Cd低积累水稻品种及实际生产应用中,应当将土壤Mn含量纳入品种适用性的考虑范围. ...
利用CRISPR/Cas9敲除OsNramp5基因创制低镉籼稻
5
2019
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
... 随着消费水平的提高,人们对稻米品质的要求也越来越高,安全、健康营养的稻米已成为最基本的需求.Cd富集在水稻中会影响稻米的品质,经人体摄入后会影响健康.有研究表明OsNramp5变异几乎对米质没什么影响,在该研究中突变体的垩白粒率和垩白度小幅下降,蛋白含量轻微增加,而且在一些品种中直链淀粉含量小幅下降,几乎不影响稻米的食味品质[49].Wang等[20]研究表明OsNramp5突变对籽粒中糙米率、精米率、直链淀粉含量、胶稠度和蛋白质含量没有显著影响.随着OsNramp5突变程度的增加,整精米率显著降低,垩白度和垩白粒率显著增加.通过改变OsNramp5的突变位点可以来缓解垩白度和垩白粒率等品质性状.董家瑜等[58]研究表明在正常Mn浓度的大田条件下,以华占为背景的osnramp5突变体的稻米品质较野生型均无显著差异,但以中花11为背景的突变体的胶稠度降低;且开花期高温造成以中花11为背景的的osnramp5突变体的垩白粒率和垩白度上升;在低Mn浓度大田条件下,2种遗传背景的osnramp5突变体的垩白粒率和垩白度上升.在水稻中,OsNramp5同时影响Cd和Mn的吸收和转运,OsNramp5变异降低水稻植株中Cd积累量的同时,也会降低Mn的含量.研究结果显示,高Mn环境可以一定程度改善由OsNramp5变异导致的米质变劣问题[58].因此,在后续利用OsNramp5基因变异选育Cd低积累水稻品种及实际生产应用中,应当将土壤Mn含量纳入品种适用性的考虑范围. ...
利用CRISPR/Cas9技术创制镉低积累香型水稻
1
2021
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
利用CRISPR/Cas9技术创制镉低积累香型水稻
1
2021
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
Large-scale profiling of brown rice ionome in an ethyl methanesulphonate-mutagenized hitomebore population and identification of high- and low-cadmium lines
2
2016
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
Gene identification and transcriptome analysis of low cadmium accumulation rice mutant (lcd1) in response to cadmium stress using MutMap and RNA-seq
3
2019
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
Knockout of OsNRAMP5 enhances rice tolerance to cadmium toxicity in response to varying external cadmium concentrations via distinct mechanisms
1
2022
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport
1
2012
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
1
2019
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
1
2019
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
Rice reduces Mn uptake in response to Mn stress
1
2018
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
OsNRAMP1 transporter contributes to cadmium and manganese uptake in rice
2
2020
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... OsNramp5是水稻根中Cd吸收与转运的蛋白,同时也是Mn的转运蛋白.Mn是植物必需的微量金属元素,与许多酶的活性有关,是多种酶的成分和活化剂,能促进碳水化合物代谢和氮的代谢,与作物生长发育和产量有关.Mn与水稻的光合作用、呼吸作用以及硝酸还原作用都有密切的关系.缺Mn时,水稻的光合作用明显受到抑制.Mn能加速水稻萌发和成熟,增加磷(P)和钙(Ca)的有效性.许多研究表明,敲除OsNramp5不单单是降低Cd积累,也会降低水稻对Mn的吸收及转运,从而对植株生长发育产生影响.缺Mn的水稻叶片叶脉间断失绿,出现棕褐色小斑点,严重时斑点连成条状,扩大成斑块[56].在低Mn条件下,OsNramp5敲除系地上部及根系的生长受到抑制,叶片出现严重的失绿症状,而随着Mn供应量的增加,苗期生长抑制表型被部分逆转,证明低Mn条件下OsNramp5基因敲除系生长受阻是由缺Mn引起的[40,43].而在Ishikawa等[41]的研究中,野生型和突变体lcd-kmt1及lcd-kmt2的植株生长几乎没有差异.但水培试验中,在Mn充足的情况下,可能是由于突变类型的不同导致其表型有差异,lcd-kmt3的生长受到抑制,株型小,且抽穗时间明显早于野生型.Tang等[46]研究显示,突变体的地上部和根部Cd和Mn含量均显著低于野生型.在低Mn条件下,突变体表现出严重的生长发育迟缓和叶片变黄.然而,在2 μmol/L或更高的Mn浓度下,突变体的生长与野生型类似.Yang等[47]发现在低Mn条件下,与野生型植株相比,OsNramp5突变体除了根和地上部生长受到严重抑制外,还表现出叶绿素含量降低和黄化症状.而随着Mn浓度的增加,植株生长和叶绿素含量逐渐恢复.并且当Mn浓度达到12.5 mol/L时,缺陷生长和叶绿素缺乏表型在突变体中得到完全拯救.龙起樟等[49]发现OsNramp5基因敲除材料的株高小幅度降低.Chang等[57]将OsNramp5与OsNramp1进行双基因敲除,在缺Mn处理中突变体发育迟缓甚至死亡.另有研究发现OsNramp5编码区不同位置的敲除对水稻植株生长可能产生不同影响.敲除位点在基因第7外显子的植株生长正常,而敲除位点在第9外显子的植株生长严重受抑、植株变矮[48].另外,随着OsNramp5突变程度的升高,对植株的生长情况影响也越大[20].值得注意的是,过表达OsNramp5则对水稻植株的生长发育几乎不产生影响[21]. ...
不同锰浓度环境下OsNRAMP5突变对水稻耐热性和主要经济性状的影响
5
2021
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
58]
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
... 随着消费水平的提高,人们对稻米品质的要求也越来越高,安全、健康营养的稻米已成为最基本的需求.Cd富集在水稻中会影响稻米的品质,经人体摄入后会影响健康.有研究表明OsNramp5变异几乎对米质没什么影响,在该研究中突变体的垩白粒率和垩白度小幅下降,蛋白含量轻微增加,而且在一些品种中直链淀粉含量小幅下降,几乎不影响稻米的食味品质[49].Wang等[20]研究表明OsNramp5突变对籽粒中糙米率、精米率、直链淀粉含量、胶稠度和蛋白质含量没有显著影响.随着OsNramp5突变程度的增加,整精米率显著降低,垩白度和垩白粒率显著增加.通过改变OsNramp5的突变位点可以来缓解垩白度和垩白粒率等品质性状.董家瑜等[58]研究表明在正常Mn浓度的大田条件下,以华占为背景的osnramp5突变体的稻米品质较野生型均无显著差异,但以中花11为背景的突变体的胶稠度降低;且开花期高温造成以中花11为背景的的osnramp5突变体的垩白粒率和垩白度上升;在低Mn浓度大田条件下,2种遗传背景的osnramp5突变体的垩白粒率和垩白度上升.在水稻中,OsNramp5同时影响Cd和Mn的吸收和转运,OsNramp5变异降低水稻植株中Cd积累量的同时,也会降低Mn的含量.研究结果显示,高Mn环境可以一定程度改善由OsNramp5变异导致的米质变劣问题[58].因此,在后续利用OsNramp5基因变异选育Cd低积累水稻品种及实际生产应用中,应当将土壤Mn含量纳入品种适用性的考虑范围. ...
... [58].因此,在后续利用OsNramp5基因变异选育Cd低积累水稻品种及实际生产应用中,应当将土壤Mn含量纳入品种适用性的考虑范围. ...
不同锰浓度环境下OsNRAMP5突变对水稻耐热性和主要经济性状的影响
5
2021
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
58]
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... OsNramp5基因变异可显著降低水稻籽粒Cd含量,但对水稻产量性状的影响也同样值得关注.到目前为止,不同研究的结果不尽相同.Sasaki等[40]发现敲除OsNramp5基因会导致产量严重下降,仅为野生型的11%.Ishikawa等[41]发现,当在Cd污染的稻田中生长时,突变体lcd-kmt1、lcd-kmt2与野生型的产量无显著差异,而突变体lcd-kmt3的产量低于野生型.Yang等[47]对两个粳稻品种进行OsNramp5基因敲除,突变体的株高、结实率和每穗粒数等农艺性状都有一定程度的降低,最终导致籽粒产量略有下降,突变体植株生长的减慢可以归因于Mn含量的显著减少.作者认为在评估osnramp5突变体的适用性时,需要考虑稻田的土壤pH和土壤含水量,因为它们可能会影响土壤中有效Mn的水平,从而决定突变对粮食产量的影响.Tang等[46]利用华占基因敲除品系及其与隆科638S基因敲除品系配制获得的杂种评价了OsNramp5突变对农艺性状的影响,结果显示敲除OsNramp5不影响水稻产量.Liu等[48]研究中,敲除位点在OsNramp5基因第7外显子,植株正常生长且产量不受影响,而敲除位点在第9外显子时植株的产量显著降低,仅为野生型的55.7%.在Wang等[20]研究中,产量随着OsNramp5突变程度的降低而增加,但分蘖数降低.董家瑜等[58]研究发现华占、中花11这2种不同遗传背景、同一位点突变的osnramp5突变株系即使在相同的种植条件下,产量表现也较野生型不同,说明遗传背景对osnramp5突变株系的环境适应性影响很大.在龙起樟等[49]研究中,OsNramp5敲除株系有效分蘖略微增加,结实率和千粒重小幅降低,产量平均减少6.9%.说明对OsNramp5进行基因敲除可以显著降低籽粒Cd积累,但在某些种植条件下,代价为小幅产量损失.因此,OsNramp5功能缺失突变体产量性状表现不一,表现为产量减少或不受影响的主要原因可归结于基因变异类型的不同,同时与遗传背景及种植条件也有一定关系. ...
... 随着消费水平的提高,人们对稻米品质的要求也越来越高,安全、健康营养的稻米已成为最基本的需求.Cd富集在水稻中会影响稻米的品质,经人体摄入后会影响健康.有研究表明OsNramp5变异几乎对米质没什么影响,在该研究中突变体的垩白粒率和垩白度小幅下降,蛋白含量轻微增加,而且在一些品种中直链淀粉含量小幅下降,几乎不影响稻米的食味品质[49].Wang等[20]研究表明OsNramp5突变对籽粒中糙米率、精米率、直链淀粉含量、胶稠度和蛋白质含量没有显著影响.随着OsNramp5突变程度的增加,整精米率显著降低,垩白度和垩白粒率显著增加.通过改变OsNramp5的突变位点可以来缓解垩白度和垩白粒率等品质性状.董家瑜等[58]研究表明在正常Mn浓度的大田条件下,以华占为背景的osnramp5突变体的稻米品质较野生型均无显著差异,但以中花11为背景的突变体的胶稠度降低;且开花期高温造成以中花11为背景的的osnramp5突变体的垩白粒率和垩白度上升;在低Mn浓度大田条件下,2种遗传背景的osnramp5突变体的垩白粒率和垩白度上升.在水稻中,OsNramp5同时影响Cd和Mn的吸收和转运,OsNramp5变异降低水稻植株中Cd积累量的同时,也会降低Mn的含量.研究结果显示,高Mn环境可以一定程度改善由OsNramp5变异导致的米质变劣问题[58].因此,在后续利用OsNramp5基因变异选育Cd低积累水稻品种及实际生产应用中,应当将土壤Mn含量纳入品种适用性的考虑范围. ...
... [58].因此,在后续利用OsNramp5基因变异选育Cd低积累水稻品种及实际生产应用中,应当将土壤Mn含量纳入品种适用性的考虑范围. ...
Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice
2
2014
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
... ,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
Producing cadmium-free Indica rice by overexpressing OsHMA3
1
2019
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
Resequencing of 1,143 indica rice accessions reveals important genetic variations and different heterosis patterns
2
2020
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
水稻籽粒镉低积累资源挖掘及其新材料创制
2
2021
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
水稻籽粒镉低积累资源挖掘及其新材料创制
2
2021
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
M_1TDS技术及镉低积累杂交水稻亲本创制与组合选育
4
2022
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
63]
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...
M_1TDS技术及镉低积累杂交水稻亲本创制与组合选育
4
2022
... Effects of different mutation types of OsNramp5 on metal content and agronomic traits of rice.
Table 2. 变异位置
Mutation site | 变异来源Mutation method | 遗传背景Genetic background | 变异类型
Mutation type | 籽粒Cd、Mn含量Contents of Cd and Mn in grains | 其他金属含量
Other metal content | 产量及其他性状
Yield and other traits | 参考文献Reference |
| 第1外显子 | CRISPR/Cas9技术 | 南粳46、
淮稻5号 | 1 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低,穗数增加 | [47] |
| 17 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 11 bp插入 | Cd、Mn显著下降 | 对Fe无显著影响 | 产量、株高、每穗粒数、结实率均显著降低 | [47] |
| 中花11 | 4 bp缺失 | Cd、Mn显著下降 | — | 生长严重受阻,根及地上部干质量显著降低 | [57] |
| 第2外显子 | CRISPR/Cas9技术 | 黄华占 | 1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣,分蘖数增加 | [20] |
| 第6外显子 | CRISPR/Cas9技术 | 黄华占 | 2 bp插入 | Cd、Mn显著下降 | 对Fe、Zn无显著影响 | 产量、每穗粒数、结实率、秸秆产量均显著降低,米质变劣 | [20] |
| 第7外显子 | CRISPR/Cas9技术 | 锡稻1号 | 1 bp插入 | Cd显著下降 | — | 产量、其他性状无显著影响 | [48] |
| EMS诱变 | 9311 | SNP变异 | Cd显著下降 | 对Fe、Zn、Cu无显著影响 | 其他性状无显著影响 | [52] |
| 第8外显子 | EMS诱变 | Hitomebore | SNP变异 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [51] |
| 第9外显子 | 碳离子束辐射诱变 | 越光 | 1 bp缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn、Cu无显著
影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占
| 3 bp缺失+
1 bp插入 | Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 5 bp缺失
| Cd<0.05 mg/kg,
Mn极显著下降 | 对Cu、Zn无显著影响,
Fe显著上升 | 产量、其他性状无显著影响 | [46] |
| 锡稻1号 | 33 bp缺失 | Cd显著下降 | — | 减产44.3%,生长严重受阻,株高降低 | [48] |
| 黄华占 | 1 bp插入 | 均显著下降 | 对Fe、Zn无显著
影响 | 产量、每穗粒数、结实率、秸秆生物量均显著降低 | [20] |
| 中花11 | 2 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 中花11 | 5 bp缺失+
1 bp插入 | — | — | 产量、千粒重极显著降低,米质变劣 | [58] |
| 第10外显子 | 碳离子束辐射诱变 | 越光 | 433 bp插入 | Cd<0.05 mg/kg,Mn显著下降 | 对Fe、Zn、Cu
无显著影响 | 产量、其他性状无显著影响 | [41] |
| CRISPR/Cas9技术 | 华占、
五丰B、
五山丝苗、
中早35 | 1-3 bp缺失+
1 bp插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu、Ca、
As、Se无显著影响 | 减产6.9%,株高、结实率、千粒重小幅降低,有效分蘖略微增加 | [49] |
| 第5内含子 | T-DNA插入 | 中花11 | 大片段插入 | — | 对K、Ca、Mg、Zn、
Cu无显著影响 | 生长受阻,叶片变黄 | [43] |
| 第8内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 18 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第10内含子 | 碳离子束辐射诱变 | 隆臻36S、
华恢8612 | 3 bp缺失 | Cd、Mn显著下降 | — | 产量、其他性状无显著影响 | [63] |
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... [
63]
| 第12内含子 | T-DNA插入 | 中花11 | 大片段插入 | Cd、Mn显著下降 | 对Fe、Zn、Cu无
显著影响 | 减产89%,生长受阻,叶片严重失绿 | [40] |
| 全基因缺失 | 碳离子束辐射诱变 | 越光 | 227 kb缺失 | Cd<0.05 mg/kg,
Mn显著下降 | 对Fe、Zn无显著影响,
Cu显著上升 | 产量显著降低,抽穗早,株型小,穗数多,但秸秆产量低 | [41] |
| 全基因缺失 | 60Co辐射诱变 | 粤泰B | 408 kb缺失 | Cd显著下降 | — | — | [61-62] |
“—”表示数据不详. ...
... 水稻籽粒中Cd的积累主要经历3个过程:根转运体从根际土壤吸收Cd2+进入根细胞,木质部装载介导Cd2+从根转移到茎,Cd2+经过韧皮部从茎转运到籽粒[44].OsNramp5在水稻对Cd的吸收和转运中都发挥着重要的作用.Sasaki等[40]针对粳稻品种中花11的OsNramp5第12内含子T-DNA插入突变体进行研究,结果表明突变体的根部和地上部的Mn、Cd含量均显著低于野生型,Zn、Cu含量差异不显著.将野生型及敲除系的幼苗在25 ℃和4 ℃下用不同浓度的Cd或Mn溶液处理30 min,测定根部对Cd和Mn的吸收,结果显示突变体几乎丧失了吸收Cd和Mn的能力.Ishikawa等[41]在粳稻越光背景下利用碳离子束辐射诱变获得了3个水稻osnramp5突变体(osnramp5-1、osnramp5-2 和 osnramp5-3),其中osnramp5-1为第10外显子433 bp插入,osnramp5-2为第9外显子缺失1 bp,osnramp5-3为整个OsNramp5缺失.研究表明,突变体编码的缺陷转运蛋白极大地降低了根系对Cd的吸收,导致茎叶和籽粒中Cd的减少,而Fe、Zn、Cu的含量没有显著差异.这三个突变体降低Cd吸收的同时也降低了对Mn的吸收,这说明Cd可能是通过Mn的根系吸收途径被吸收,但其他矿质营养元素的吸收没有受到影响.当在Cd污染的稻田中生长时,突变体的籽粒中几乎检测不到Cd,并且没有表现出不利的农艺或经济性状.Yang等[43,45]从水稻突变数据库(RMD)中获得了一个中花11 OsNramp5第5内含子T-DNA插入突变体osnramp5,OsNramp5完全不表达,该突变体在根和地上部的Cd浓度显著降低,且减少了Mn从根向地上部的运输,导致地上部Mn无法累积.较高的Mn供应可以弥补突变体根系吸收能力的丧失,但不能弥补Mn从根向地上部的转运.野生型对Cd的吸收与Mn的施用量之间呈现显著的负相关,而突变体osnramp5不存在这样的趋势.笔者认为水稻植株对Mn和Cd的吸收存在拮抗关系,且该拮抗由OsNramp5介导.Tang等[46]利用CRISPR/Cas9基因编辑技术对籼稻华占和隆科638S的OsNramp5第9外显子进行基因敲除,突变类型鉴定结果显示除1个突变体是跨越两个靶点的225 bp DNA片段缺失类型外,其余均为短插入或短缺失(InDel).水培试验表明osnramp5突变体的地上部和根部的Cd和Mn含量均显著低于野生型.在高Cd处理条件下,野生型的生长受到严重抑制,而突变体的生长未受影响.Cd污染大田试验进一步表明,基因敲除水稻籽粒中Cd含量始终低于0.05 mg/kg,远远低于0.2 mg/kg的国家标准,为培育籽粒Cd低积累的籼稻品种提供了一条切实可行的途径.Yang等[47]选用两个粳稻品种南粳46和淮稻5号对OsNramp5进行基因编辑,在OsNramp5的第1外显子上分别有1 bp、17 bp和11 bp的缺失,得到3个移码突变的osnramp5突变体,突变体剑叶和籽粒中的Cd含量显著降低.另外,在有Mn条件下,突变体的根和地上部的Mn浓度显著低于野生型,并且两者的Mn浓度差异随着Mn供应量的增加而变大,但改变外源Mn浓度对Cd的积累无明显影响.而低Mn供应时突变体的根中Fe含量增加.Liu等[48]利用CRISPR/Cas9基因编辑技术敲除粳稻锡稻1号OsNramp5获得两个分别为第7外显子上插入1 bp,第9外显子上缺失33 bp的突变株系.Cd污染田间试验及盆栽高Cd处理试验显示,突变体均表现出较低的籽粒Cd含量(<0.06 mg/kg).Wang等[20]以籼稻品种黄华占作为受体材料进行OsNramp5基因敲除,获得转基因植株LCH1、LCH2和LCH3.其中LCH1为第2外显子上插入T,产生移码突变,导致Nramp结构域完全缺失;LCH2为第6外显子上插入AA,编码一个由100个氨基酸残基组成的Nramp结构域,LCH3为第9外显子上插入G,编码一个有202个氨基酸残基的Nramp结构域.研究表明,随着OsNramp5突变程度的降低,其对绿色组织Mn积累的影响逐渐减弱,OsNramp5的功能缺失可以降低籽粒中Cd和Mn的含量,Cd和Mn的积累趋势一致.龙起樟等[49]选取华占和五丰B以及五山丝苗和中早35为材料,通过CRISPR/Cas9技术靶向OsNramp5第10外显子创制低Cd籼稻,突变类型大多为缺失1~3 bp和插入1 bp,占所有突变的66%.OsNramp5被敲除后,水稻籽粒中Cd含量大幅下降,Mn含量减少一半左右,而Cr含量增加一半以上,Fe、Zn、Ca和Cu等含量受影响较小或不受影响.胡黎明等[50]针对OsNramp5第1内含子、第2外显子和香味基因OsBadh2第3外显子设计靶点进行基因编辑,结果显示敲除OsNramp5基因可有效降低籽粒Cd的积累,且大多数突变体Mn含量也显著降低,而Fe含量会相应增高,此法可快速创制具有Cd低积累和香味的水稻种质.有研究人员从Hitomebore的EMS诱变群体中分离到地上部及籽粒Cd水平较低的突变株系1281-m,分析OsNramp5基因组序列显示,第8外显子的第242位氨基酸由组氨酸替换为谷氨酰胺,突变体的农艺性状及产量与野生型无显著差异[51].Cao等[52]利用EMS诱变籼稻品种9311获得水稻植株和籽粒Cd积累量极低的突变体lcd1.该突变体中OsNramp5第7外显子高度保守区的第236位氨基酸脯氨酸被亮氨酸替换.因此,第236―242位氨基酸所在区域附近可能和Cd转运直接相关,当SNP位点突变发生在该区域时,水稻籽粒Cd含量显著降低且对其他农艺性状无明显影响(图1).另外,突变位点发生在OsNramp5基因第8和第10内含子时,突变体的产量及其他农艺性状也无显著变化[63].Tang等[53]发现,对OsNramp5进行基因敲除有效地补充了突变体植株地上部缺乏的矿质营养,从而间接地提高了水稻对高Cd胁迫的耐受性.也有研究人员发现OsNramp5的RNAi敲低突变体的根、地上部和木质部汁液中的Mn均显著低于野生型[42,54].此外,许多研究表明OsNramp5突变对水稻中Fe、Zn、Cu等金属元素含量几乎没有影响[42-43,46 -47,55]. ...
... 目前的研究表明,通过OsNramp5基因功能缺失和组成型过量表达OsHMA3[59]是培育Cd低积累水稻品种最有效的途径.尽管过表达OsHMA3基因能显著降低Cd含量,且对Mn无影响,利用这一措施降Cd本应该是最优的选择,但该途径涉及到转基因,短时间内无法生产应用.相较于OsNramp5突变体创制,通过非转基因的方式创制OsHMA3表达量升高的种质工作难度非常大.OsNramp5基因功能缺失在显著降低Cd的同时也会影响Mn的吸收,在缺Mn的条件下会表现出不利的表型,限制了其在低Mn农田中的应用.但在我国南方一些地区Cd、Mn复合污染的情况下,利用OsNramp5基因功能缺失具有巨大的潜力.由于受我国转基因政策的限制,这些优异种质还不能用于生产.迄今只有lcd1突变体、珞红3A、珞红4A及对应的保持系以及利用重离子诱变获得的莲1S、R001和韶香100等少数几个Cd低积累资源能被直接应用[52,59⇓⇓⇓ -63].目前利用lcd1突变体、珞红3A、珞红4A转育的湘安早系列、安优系列、西子系列、DY系列等低镉水稻品系已进入湖南省农业科学院组织开展的Cd低积累水稻品种自主区域试验,其中一些材料综合表现优异,成功进入续试;而莲1S和R001分别是通过诱变隆臻36S和华恢8612获得,其配组的低Cd杂交组合莲两优1号表现出非常高的产量潜力.一般情况下,OsNramp5基因功能缺失水稻能够在中、重度Cd污染稻田中生产Cd含量达标的稻米,但其能够耐受Cd污染的浓度还没有一个明确范围,需进一步探索;同时,OsNramp5基因功能缺失导致Mn含量降低,是否会影响水稻对病、虫、草害以及高温、低温等逆境的耐受还有待深入研究. ...


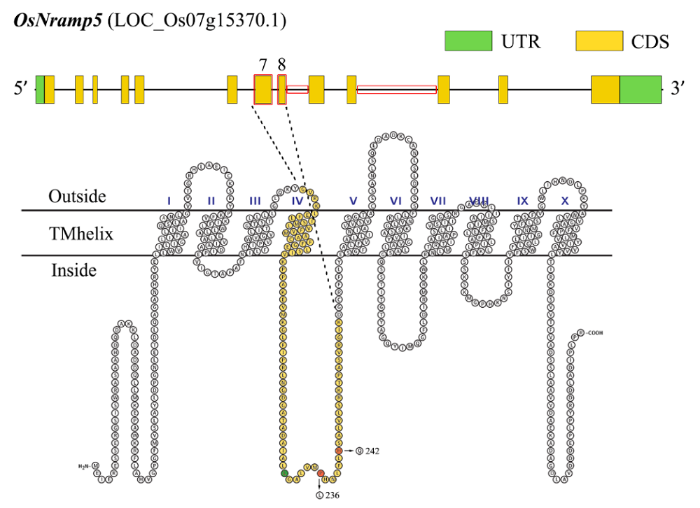

{kind=link}
{kind=link}